
摘要
:
脂质是机体内重要的营养物质之一,参与机体的能量供应、生物膜形成以及代谢调节等,内质网是其合成的主要场所。近年来越来越多研究表明,细胞色素
P450
酶的表达和活性影响机体对有害物质的敏感性以及药物的疗效,影响疾病的发生及发展,而细胞色素
P450
酶作为膜蛋白受到膜组成的影响。就细胞色素
P450
酶调控过程中内质网脂质稳态的作用进行综述。
酶是生命活动中重要的催化活性物质,参与一系列生理生化反应,决定了机体的正常生理病理状态。肝脏作为体内最大的代谢器官,含有大量的酶参与糖、脂、蛋白、维生素等的代谢,具有防御、解毒功能,其中代谢酶参与外源性异物的代谢并促进其排出体外。细胞色素
P450
(
CYP450
)酶系作为肝脏中主要的代谢酶家族,是机体内最重要的
Ⅰ
相代谢酶,广泛参与外源性药物、环境毒物、致癌物以及内源性激素等的代谢过程。研究显示,
CYP450
的表达及活性的差异将影响物质在体内的代谢,关系到药物的疗效以及疾病的进展
[
1-2
]
。
内质网是细胞内蛋白、脂质、寡糖的合成和修饰以及钙离子贮存的位点,同时也负责内源性及外源性产物的脱毒处理
[
3
]
。当机体受到如氧化还原稳态的失衡、病毒感染、钙离子浓度失调等刺激时,内质网蛋白合成功能受到干扰,导致大量未折叠及错误折叠蛋白蓄积在网腔内引发内质网应激,细胞进而通过激活未折叠蛋白反应(
unfolded protein response
,
UPR
)恢复内质网蛋白质稳态的平衡
[
4
]
,而持续的内质网应激可引发炎症、细胞凋亡,影响相关疾病病程的进展。磷脂是膜的主要组成部分,通过内质网合成和修饰以及高尔基体的运输参与构成各种生物膜
[
5
]
。线粒体是体内重要的产能细胞器,其功能和结构依赖于外膜和内膜特定的脂质组成,即高含量的非双层磷脂,当线粒体膜组成改变时将影响细胞的自噬和凋亡
[
6
]
。因此,内质网是整个细胞脂质水平的中枢调节器,影响细胞内各种生物膜的构成,可能影响细胞功能甚至增加患病的风险
[
7
]
。
CYP450
酶是真核生物中主要分布在内质网内膜上的膜蛋白,研究显示膜蛋白受到环境的影响,可以与脂质、膜骨架以及其他膜蛋白发生复杂的相互作用,导致在不同的环境中表现出不同的扩散形式
[
8
]
。本文主要探讨了内质网内脂质稳态在
CYP450
调控过程中的作用。
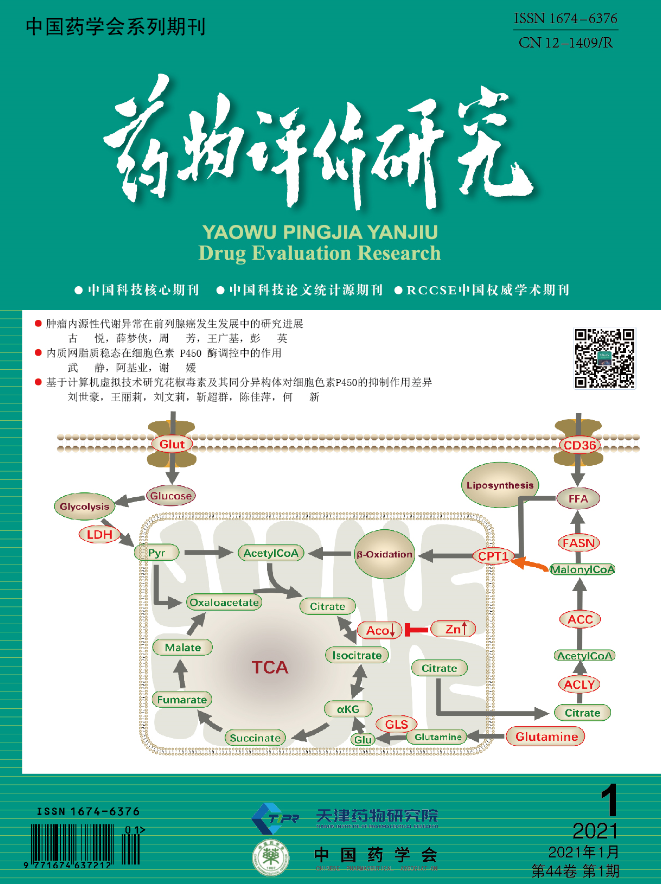
1
脂质的合成与功能
体内脂质主要包括三酰甘油、胆固醇和磷脂
3
大类,其来源主要有
2
个方面:一是通过肝脏摄取循环中来自食物或脂肪组织的游离脂肪酸;二是利用乙酰辅酶
A
进行脂质的从头合成(
denovo lipogenesis
,
DNL
)。首先,线粒体中三羧酸循环(
TCA
)产生的柠檬酸盐转运至细胞质,并在光面内质网上通过一系列裂解、转化、羧化以及合成反应生成含
16
个碳单位的棕榈酰酯,再经过去饱和酶作用产生单不饱和脂肪酸或多不饱和脂肪酸。生成的脂肪酸一部分以三酰甘油的形式存储,一部分经过修饰最终作为信号分子或成为构建生物膜体系的鞘脂和甘油磷脂,最终通过分泌途径或内质网接触位点运输到细胞内其他部位
[
9
]
。三酰甘油是体内重要的能量储备,并作为脂肪组织分布于皮下、内脏周围,起着隔热和保护机体的作用。胆固醇是组成细胞膜的重要成分,它不但确保细胞内部可以正常运作,还确保了细胞膜内外物质的流动性,以完成不间断的物质交换和新陈代谢
[
10
]
。胆固醇还是合成各类激素、维生素
D
以及胆汁酸的原料,以维持体内的信号传递和代谢
[
11-12
]
。磷脂作为各类生物膜的基本骨架,控制着细胞的新陈代谢、信号传导并发挥抵抗外部侵害、修复细胞的作用
[
13
]
。细胞的各类生命活动也离不开酶的催化,而生物膜上的许多酶与磷脂密切相关。
2
内质网脂质稳态对
CYP450
酶活性的调控研究
2.1
内质网与脂质稳态
脂质是细胞内重要的营养物质之一,饮食、运动以及外界刺激等都会引起机体内脂质的变化
[
14-16
]
,内质网是细胞内脂质合成和调节的重要场所,内质网为适应外界环境变化对体内脂质合成和降解平衡的动态调节称为内质网脂质稳态。
2.1.1
内质网相关降解对脂质的调控
内质网是脂质的重要合成位点
[
17
]
,并且作为脂质代谢的中心枢纽,感应并维持脂质稳态,包括脂质合成相关基因表达的调控、酶活性的变构调节以及翻译后修饰等。细胞内胆固醇的水平主要是通过固醇调节元件结合蛋白(
sterol regulatory element binding protein
,
SREBP
)进行调控
[
18
]
,但
SREBP
对胆固醇的调控属于转录前调控,需要消耗较长时间。
3-
羟基
-3-
甲基戊二酸单酰辅酶
A
还原酶(
HMGCR
)是胆固醇合成过程中的限速酶,可催化羟甲戊二酰辅酶(
HMG-CoA
)转化为甲羟戊酸。当体内胆固醇水平过高时,内质网通过对胆固醇合成过程中的限速酶
HMGCR
和角鲨烯单加氧酶进行泛素化标记直接快速降解,对脂质合成相关的酶进行转录后水平的调控,实现了机体内胆固醇水平的快速调控
[
19
]
,这种过程称为内质网相关降解(
ER-associateddegradation
,
ERAD
)。
2.1.2
内质网内分子伴侣蛋白对脂质的调控
脂质合成的前体来源于细胞外以及细胞质内脂滴,其转运依赖微粒体三酰甘油转移蛋白(
microsmal triglyceridetransfer protein
,
MTTP
)。
MTTP
是内质网内的一种分子伴侣蛋白,可将磷脂和三酰基甘油转移到新生的载脂蛋白
B
上组装脂蛋白。在缺乏
MTTP
的情况下,膜相关的载脂蛋白
B
发生降解,影响脂质的转移和运输,引起血浆中三酰甘油、胆固醇、鞘磷脂和神经酰胺水平的降低
[
20
]
。
2.1.3
内质网应激对脂质的调控
内质网应激是机体在应激状态下的一种重要的信号反应通路,属于细胞的自我保护机制。研究发现内质网应激激活了
cAMP/PKA
途径以及
ERK1/2
途径,进而引起下游激素敏感性脂肪酶
Ser563
和
Ser660
两个位点磷酸化以及脂滴表面
Perilipin
磷酸化,最终导致脂肪分解
[
21
]
。此外,
Yao
等
[
22
]
发现内质网应激通过上调
CD36
的表达促进脂质在巨噬细胞中的积累。内质网应激还通过过表达
miR-33
促进胆固醇的摄取、抑制胆固醇的流出以及调节相关转运蛋白的表达,调节脂质分解代谢过程
[
23
]
。
2.1.4
内质网内
Ca
2+
水平对脂质的影响
Ca
2+
是细胞内重要的信号分子,调控细胞增殖、分泌、骨骼肌收缩
[
24
]
以及死亡
[
25
]
等一系列生命活动。细胞内
Ca
2+
主要储存在内质网腔内,其余以游离形式存在于胞浆中。内质网腔内的
Ca
2+
调节内质网许多功能,包括蛋白的合成、修饰、转运以及脂质稳态等
[
26
]
。
SREBPs
超家族在内质网胆固醇合成过程中发挥重要作用,研究显示在钙调蛋白缺乏导致内质网内
Ca
2+
耗竭的细胞中,
SREBP
活性增强且不受细胞内未酯化胆固醇含量、内质网应激以及高尔基体上
S1P
和
S2P
蛋白酶活性的影响,导致细胞内胆固醇水平失调以及未酯化胆固醇的迁移
[
27
]
。此外,
Ca
2+
还能通过静电作用导致囊泡内磷脂的横向分布和相变发生改变
[
28
]
。
2.1.5
内质网和其他细胞器间相互作用参与脂质调节
内质网是细胞内膜结构较大的细胞器,内连核膜,外连细胞膜,是细胞内物质运输的重要通道。研究显示,内质网和细胞膜接触位点不仅作为脂质传递的通道,还是整合脂质合成途径的调节点,当接触位点缺乏时将影响磷脂的生物合成进而导致质膜脂质组成的改变,使得质膜中甾醇发生紊乱
[
29
]
。高尔基复合物是分泌系统的中心枢纽,接收从内质网新合成的蛋白质和脂质并进行脱羧等修饰,积极参与蛋白质和脂质的生物合成并将其运输到目的位置,当内质网和高尔基体之间接触位点受阻时将影响脂质的交换和修饰导致脂质稳态失衡
[
30
]
。此外,线粒体和内质网之间也通过线粒体
-
内质网相互作用膜结构(
mitochondrial-associatedER membrane
,
MAM
)进行物质的传递及信息的交流。由于内质网和线粒体具有不同的脂质加工酶,故内质网上合成的脂质需要线粒体膜上酶的协同作用,
MAM
缺陷也将影响磷脂以及三酰甘油的合成和转运
[
31-32
]
。
2.2
内质网脂质稳态对
CYP450
酶的调控
CYP450
酶的催化结构域与膜存在相互作用
[
33
]
,因此膜环境会影响
CYP450
酶的催化活性。磷脂作为形成脂质双层的关键组分,对于真核细胞膜的结构、功能和稳定是非常重要的,也是
CYP450
酶发挥正常功能的关键。
CYP450
氧化还原酶(
Cytochrome P450 Reductase
,
CPR
)作为真核细胞
CYP450
酶催化系统中的主要电子供体锚定在内质网上,而磷脂酰胆碱(
phosphatidylcholine
,
PC
)是内质网膜磷脂层的主要成分,也是
CYP450
单加氧酶系统的重要脂质成分,帮助实现从
CPR
到
CYP450
的最佳电子转移。因此,
PC
水平与
CYP450
和
CPR
的活性呈正相关
[
34
]
。
对哺乳动物微粒体酶活性的研究显示,阴离子膜磷脂如磷脂酸(
phosphatidic acid
,
PA
)、磷脂酰丝氨酸(
phosphatidylserine
,
PS
)和磷脂酰肌醇(
phosphatidylinositol
,
PI
)能产生负电荷接口促进
P450
嵌入膜中从而增强
CYP450
酶的活性
[
35-36
]
,体外构建的重组膜模型也显示增加膜中阴离子膜磷脂的含量可以增强
P450
酶的活性
[
37
]
。
有研究显示,膜脂质双层中的磷脂组成还会影响
CYP450
酶和底物的结合从而影响酶的活性
[
34
,
38
]
,且其组成和含量也决定了膜的通透性和流动性。膜的流动性会影响膜蛋白的构象和功能,膜上的长链多不饱和脂肪酸(
polyunsaturated fatty acid
,
PUFA
)的增加会导致膜流动性的增加,使得膜蛋白分子活性增强,进而增强了细胞以及组织的代谢活性
[
39
]
。
此外,内质网还包含不同的脂质微区(
lipid microdomains
),包括富含不饱和脂肪酸的
ld
区域(
liquiddisorder membrane
)和富含高浓度饱和脂肪酸、胆固醇以及鞘磷脂的
lo
区域(
liquidorder membrane
),其中胆固醇是质膜有序结构形成的关键,一旦胆固醇耗竭,将导致膜上结合蛋白的释放。
CPR
和
CYP450
有序地分布在膜上,其中
CPR
主要存在于
lo
区域,而不同的
CYP450
酶亚型差异性地分布在
lo
或
ld
区域,或在两区域均匀分布
[
40-41
]
。由于
CYP450
需要和
CPR
发生相互作用才能发挥催化功能,因此定位于
lo
区域有利于
CYP450
活性的发挥。而
CPR
和
CYP2E1
由于分布于不同区域导致相互作用减弱,但也限制了
CYP2E1
产生
H
2
O
2
这种损伤性活性氧簇
[
42
]
。此外,膜上胆固醇的含量也会直接影响
P450
酶的活性,在正常情况下胆固醇的存在帮助实现膜的增厚和有序化,使得
CYP3A4
更大程度的浸入膜中,但过多的胆固醇会降低
CYP3A4
膜浸入部分的柔韧性,影响底物进入酶的活性位点
[
43
]
。
脂质也通过影响
CYP450
酶的基因表达进行转录调控。
CYP4Fs
催化包括不饱和长链和支链脂肪酸、含有长链烷基侧链的维生素、白三烯、前列腺素和羟基二十碳四烯酸的
n-
羟基化反应,
Massaro
等
[
44
]
发现
n-3
多不饱和脂肪酸通过上调
CYP4Fs
基因,导致
CYP4Fs
的蛋白表达水平增加。
上述研究表明,脂质稳态在
CYP450
酶的表达及活性的调控中发挥着重要作用。
3
脂质调控
CYP450
酶对疾病进展的影响
CYP450
酶广泛参与内、外源性物质的代谢,其活性的变化与多种肝脏疾病、肿瘤以及帕金森病等密切相关。
CYP2E1
是肝脏中丰富的代谢酶之一,可以将乙醇、四氯化碳以及亚硝胺等活化为具有生理功能以外肝毒性或致癌性的物质。丙烯酰胺是人类生活中普遍存在的有毒物质,
CYP2E1
是唯一负责将丙烯酰胺转化为高反应性代谢产物缩水甘油酰胺的酶,后者与
DNA
形成加合物。
Katen
等
[
45
]
研究表明精母细胞中的
CYP2E1
代谢活化丙烯酰胺引发精子
DNA
损伤导致胚胎致死率的升高,
Guo
等
[
46
]
还发现高催化活性的
CYP2E1
通过限制巨噬细胞向
M2
表型的极化加剧了肝纤维化进程。非酯化脂肪酸(
nonesterifiedfattyacid
,
NEFA
)是脂质的一种,通常在血中浓度很低,其含量水平极易受脂代谢、糖代谢和内分泌机能等因素影响。研究显示体内高水平的
NEFA
会引起细胞脂毒性,导致肝中
CYP2E1
的表达和活性被过度诱导。
CYP2E1
的过表达导致
ROS
过量产生并引起氧化损伤,过量的
ROS
又会引起脂质过氧化,蛋白质结构变化以及核酸的链断裂,引起细胞膜完整性和功能的改变来破坏细胞和细胞器,从而引发
“
二次打击
”
并导致细胞凋亡以及疾病
[
47
]









