

摘 要:
蟾皮为蟾蜍科动物中华大蟾蜍
Bufo bufo
gargarizans
或黑眶蟾蜍
B. melanosticus
的干燥全皮,已纳入壮药名录。作为一种民族药,常用于满族、回族和朝鲜族的民间药方,具有清热解毒和利水消肿之功效。目前已从蟾皮中发现
100
多个化合物,包括蟾蜍二烯羟酸内酯类、吲哚生物碱类、蟾蜍环酰胺类以及甾体类等成分。随着国内外药理学研究的不断深入,蟾皮单体成分呈现诸多药理作用,如抗肿瘤、抗炎、免疫调节、抗乙肝病毒、强心等,并且对特异的药物代谢酶具有较高的敏感性。通过查阅近
20
年来的国内外文献,对蟾皮的化学成分、药理活性、单体成分与药物代谢酶的相互作用等方面进行综述,以期为蟾皮更好的开发和应用提供参考。

蟾皮,别名癞蟆皮、蟾蜍皮、干蟾皮、蛤蚆皮,状语称“能唝酬”,最早记载于《本经逢原》,为蟾蜍科动物中华大蟾蜍
Bufo bufo
gargarizans Cantor
或黑眶蟾蜍
B. melanosticus
Schneider
的干燥全皮,辛,凉,微毒,经方入药,目的多为以毒攻毒
[1-2]
。《中药大辞典》记载“蟾皮能清热解毒、利水消胀,
治疗痈疽、肿毒、瘰疬、肿瘤、疳积腹胀和慢性气管”
[3]
。蟾皮作为一种民族药,已纳入壮药名录,是广西常用特色壮、瑶药材,民间广泛用于治疗恶性肿瘤
[2]
。满族常用蟾蜍细末外敷治疗疔毒疮和臁
疮腿
[4]
;回族则用干蟾蜍对急性炎症进行止痛消肿
[5]
;
而朝鲜族直接取蟾蜍皮烧熏治疗肠头挺出
[6]
。蟾皮及其制品临床应用较为广泛,特别是以干蟾皮为主要原料制成的水溶性制剂华蟾素为我国自行研发的二类新药,对原发性肝癌、肺癌、食管癌、结肠癌、乳腺癌等恶性肿瘤具有非常好的疗效
[7]
。目前已从蟾皮中分离获得
100
多个单体化合物,主要是蟾蜍二烯羟酸内酯类、吲哚生物碱类、蟾蜍环酰胺类、小分子环肽类以及甾体类等成分。随着蟾皮有效成分研究的不断深入,其中的单体成分在抗肿瘤、抗炎、免疫调节和强心等方面有显著药理活性。本文对蟾皮中化学成分和药理活性的研究进展进行梳理,为进一步开发和利用蟾皮提供理论依据和文献支持。
1
蟾皮的化学成分
蟾皮中化学成分复杂,通过分离与鉴定,已发现其中包含有蟾蜍二烯羟酸内酯类、吲哚生物碱类、蟾蜍环酰胺和小分子环肽类、甾体及其他类化合物
[8-9]
。
1.1
蟾蜍二烯羟酸内酯类化合物
此类化合物属于强心甾体类化合物,以游离型或与硫酸盐、二羧酸酯和氨基酸在
C-3
位结合的形式存在,其结构特点是
C-17
位连有一个
α-
吡喃酮基团
[10]
。根据是否与有机酸相连可分为蟾蜍毒素类和蟾毒配基类,其中蟾蜍毒素类化合物只存在于新鲜的蟾皮中,经干燥加工,蟾蜍毒素类化合物的
3
位酯键完全水解,得到相应的蟾毒配基,部分水解则形成丁二酸(辛二酸等)蟾毒配基单酯
[8]
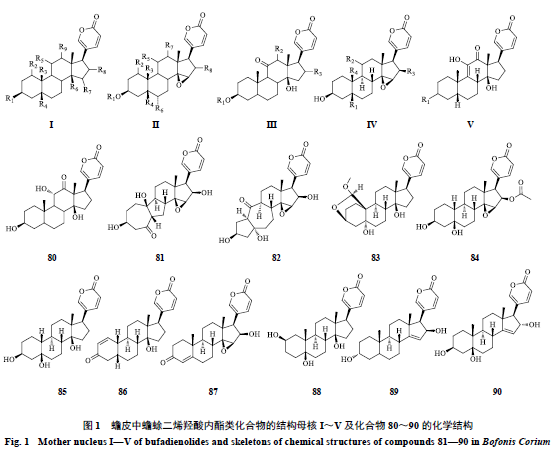
。目前,从蟾皮中已分离得到蟾蜍二烯羟酸内酯类化合物
90
个,根据母核上取代基的不同,大致分为
5
类,母核结构见图
1
(
I
~
V
为母核,化合物
80
~
90
为无法归纳入母核的化合物),分离得到的化合物见表
1
。



1.2
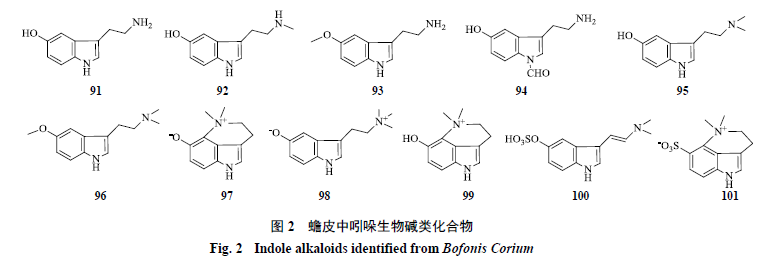
吲哚生物碱类化合物
此类化合物含有吲哚环,是神经递质
5-
羟色胺及其代谢产物,目前已从蟾皮中分离获得
11
个吲哚生物碱类化合物,分别是
5-
羟色胺(
serotonin
,
91
)
[23]
、
N
-methyl serotonin
(
92
)
[24]
、
O
-methyl serotonin
(
93
)
[24]
、
N
′-formyl serotonin
(
94
)
[24]
、蟾毒色胺(
bufotenine
,
95
)
[20]
、
5-methoxy bufotenine
(
96
)
[20]
、脱氢蟾蜍色胺(
dehydrobufotenine
,
97
)
[23]
、蟾蜍色胺内盐(
bufotenidine
,
98
)
[24]
、去氢蟾蜍色胺氢溴酸盐(
dehydrobufoteininehydrobromide
,
99
)
[25]
、蟾蜍噻咛(
bufothionine
,
100
)
[23-24]
、脱氢蟾蜍噻咛(
dehydrobufothionine
,
101
)
[26]
,结构见图
2
。
1.3
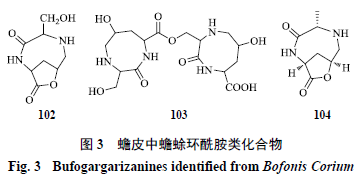
蟾蜍环酰胺类和小分子环肽类化合物
有研究者从蟾皮中分离得到蟾蜍环酰胺类化合物,包括含有内酯结构化合物,如蟾蜍环酰胺
B
(
bufogargarizanineB
,
102
)
[25]
和蟾蜍环酰胺
D
(
bufogargarizanineD
,
104
)
[26]
以及不含内酯结构化
合物,如蟾蜍环酰胺
C
(
bufogargarizanine C
,
103
)
[25]
(图
3
);此外,还分离获得环(脯氨酸
-
甘氨酸)二肽
[cyclo(Pro-Gly)dipeptide]
[23]
和环(丙氨酸
-
丙氨酸)二肽
[cyclo(Ala-Ala)dipeptide]
[26]
等小分子环肽类化合物。
1.4
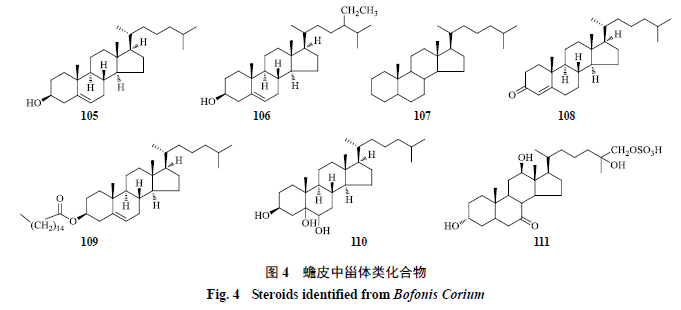
甾体类
蟾皮中已分离得到的甾体类化合物有胆甾醇
(
cholesterol
,
105
)
[9,27]
、
β-
谷甾醇(
β-sitosterol
,
106
)
[27]
、
胆甾烯醇(
cholestenol
,
107
)
[11]
、胆甾烯酮(
cholestenone
,
108
)
[26]
、棕榈酸胆甾烯酯(
palmitic acid cholesteryl ester
,
109
)
[9,27]
、
Cholestane-3β,5α,6β- triol
(
110
)
[9]
、
3α,12β,25,26-tetrahydroxy-7-oxo-5β- cholestane 26-
O
-sulfate
(
111
)
[14]
,结构见图
4
。
1.5
其他类
蟾皮中还含有腺苷、嘌呤类、嘧啶类、氨基酸、有机酸、光色素等
[9,23,24,26,28]
。此外,徐乃玉等
[29]
在中华大蟾蜍皮粉中测得钙、镁、钠、锰、铁、锌、铜、磷、硅及银等元素,其中以钙最多,其次是铁和镁。
2
蟾皮的药理活性
《中药大辞典》记载“蟾皮能清热解毒、利水消胀,治痈疽、肿毒、瘰疬、肿瘤、疳积腹胀和慢性气管”
[3]
。现代医药学研究表明,蟾皮具有抗肿瘤、抗乙肝病毒等药理作用,临床上主要用于治疗乙型
肝炎、慢性支气管炎、喉咙肿痛、痈肿疔毒等病症
[27]
;
还可用于治疗顽固性皮肤病,如带状疱疹、荨麻疹、黄褐斑、银屑病等
[30]
。目前上市产品如华蟾素片(口服液、注射液)、华蟾素滴丸、鹤蟾片、季德胜蛇药片等均含有蟾皮成分
[31]
。
2.1
抗肿瘤作用
研究表明,蟾蜍二烯羟酸内酯类化合物是蟾皮中具有抗肿瘤活性的主要成分
[32]
,能抑制肿瘤细胞增殖、诱导肿瘤细胞凋亡、抑制肿瘤细胞侵袭和转移、抗血管生成、诱导肿瘤细胞分化、逆转肿瘤细胞多药耐药等
[10]
。
2.1.1
抑制肿瘤细胞增殖
细胞增殖在肿瘤发生发展中起着至关重要的作用,因此控制细胞增殖对预防癌症至关重要
[33]
。华蟾毒精、海蟾蜍精、蟾毒灵和酯蟾毒配基对多种肿瘤细胞具有抗增殖活性,如
多发性骨髓瘤、黑色素瘤、白血病、肝癌、卵巢癌、食管鳞癌、肺癌、结肠癌等
[34-37]
。这类化合物主要是通过调控细胞周期相关蛋白,引起细胞周期阻滞,进而抑制肿瘤细胞增殖,但不同的化合物在不同肿瘤细胞中引起的细胞周期阻滞时期不同,如蟾毒灵将食管鳞癌细胞周期阻滞于
S
和
G
2
/M
期
[36]
;酯蟾
毒配基下调细胞周期蛋白
D1
(
cyclinD1
)和
cyclin E
蛋白,降低
RB
蛋白磷酸化,引起结肠癌和肺癌细胞
G
0
/G
1
期阻滞
[37]
。
ψ-bufarenogin
通过
G
2
/M
期阻滞抑制肝癌细胞增殖,下调
cyclin E
和
ki67
,上调
cyclin B1
[38]
。沙蟾毒精对食管鳞状细胞癌细胞的抗癌效果优于蟾毒灵,且对人正常的食管鳞状
Het-1A
细胞表现出更低的毒性,还可通过激活
p53
信号通路有效抑制裸鼠移植瘤生长
[39]
。与酯蟾毒配基作用类似,蟾蜍噻咛亦能抑制人肝癌细胞增殖,促进小鼠肝肿瘤
H22
细胞坏死并抑制肿瘤生长,抑制率
65.16%
,通过调控线粒体介导的凋亡蛋白抑制肝脏肿瘤生长并保护肝脏免受急性损伤
[40]
。
2.1.2
诱导肿瘤细胞凋亡
蟾皮中的有效化学成分作用于肿瘤细胞后,可转化成细胞凋亡信号,通过抑制或激活细胞内和细胞外的多种信号转导途径,诱导肿瘤细胞凋亡
[33]
。蟾毒灵促进活性氧积累,上调凋亡酶激活因子(
apoptotic protease activating factor-1
,
Apaf-1
)、
cleaved polyADP-ribose polymerase
(
PARP
)、剪切型半胱氨酸蛋白酶
3
(
cleaved caspase-3
)、
cleaved caspase-9
和
Bax/Bcl-2
,靶向线粒体依赖的信号通路诱导骨肉瘤
MG-63
细胞发生凋亡
[41]
。华蟾毒精激活
ROS/MAPKs
信号通路诱导卵巢癌细胞
[42]
和多发性骨髓瘤
U266
细胞
[34]
发生凋亡。沙蟾毒精显著上调半胱氨酸蛋白酶
(
caspase
)
-3
、半胱氨酸蛋白酶
-8
和半胱氨酸蛋白酶
-9
,
通过内源性和外源性途径激活
caspase
,有效诱导食
管鳞状细胞癌细胞发生凋亡
[39]
。
ψ-bufarenogin
抑制受体酪氨酸激酶介导的信号转导通路抑制肝癌生长
[38]
。
海蟾蜍精(
1.25 μmol/L
)引起白血病
HL-60
细胞染色质凝结、核碎裂、核溶解、细胞收缩和膨胀、胞质空泡的发生以及膜解体;降低细胞膜的完整性,导致
DNA
断裂
[35]
。在胶质母细胞瘤
U87
和
U373
细胞中,蟾蜍噻咛通过上调半胱氨酸蛋白酶
-12
、
C/EBPhomologous protein
(
CHOP
)、磷酸化蛋白激酶受体样内质网激酶(
phospho-protein kinase R-like endoplasmic reticulum kinase
,
p-PERK
)、磷酸化真核翻译起始因子
2α
(
phospho-eukaryotic translationinitiation factor 2α
,
p-eIF2α
)和转录激活因子
6
(
activatingtranscription factor 6
,
ATF6
)蛋白表达,触发内质网应激诱导细胞凋亡
[43]
。
2.1.3
抑制肿瘤细胞侵袭与转移
肿瘤细胞侵袭转移是恶性肿瘤的主要特征,是一个复杂的、多因素调控的动态过程。研究发现,蟾皮中的有效成分可通过调控相关蛋白因子抑制肿瘤细胞侵袭转移。蟾毒灵抑制活化的
B
细胞的
nuclearfactor-κ-light- chain-enhancer
和基质金属蛋白酶(
matrix metalloproteinase
,
MMP
)
-2/MMP-9
通路抑制肝癌
SK-Hep1
细胞迁移和侵袭
[44]
。沙蟾毒精下调
β-catenin
通路抑制上皮
-
间质转化,进而抑制前列腺癌
PC3
细胞的侵袭转移
[45]
。
Han
等
[46]
研究表明,酯蟾毒配基显著降低结直肠癌
HCT116
细胞侵袭和迁移能力;上调上皮标志物紧密连接蛋白和
E-
钙粘蛋白的表达,并下调纤连蛋白、波形蛋白和蜗牛蛋白的表达来破坏上皮
-
间质转化,以一种
RIP3
相关的方式抑制结直肠癌细胞侵袭和转移,显著减少小鼠脾移植
MC38
细胞产生的肝转移灶的数量和大小,最终抑制肝转移。
2.1.4
抗血管生成
血管生成被认为是癌症发展和生长的关键,发现和开发有效的血管生成抑制剂是癌症治疗的重要组成部分
[33]
。沙蟾毒精抑制血管内皮生长因子(
vascular endothelial growth factor
,
VEGF
)诱导的人脐静脉内皮细胞的增殖、迁移、侵袭、血管形成和血管内皮生长因子受体
2
(
vascularendothelial growth factor receptor 2
,
VEGFR-2
)磷酸化,通过抑制
VEGFR-2
信号通路抑制
VEGF
介导的血管生成
[47]
。华蟾毒精在体内下调血管生成抑制结肠癌的生长,以剂量相关方式显著降低人脐静脉内皮细胞分层的新血管系统和破坏血管网络的形成,减少低氧诱导因子
1α
(
hypoxia-induciblefactor 1α
,
HIF-1α
)的表达,诱导活性氧积累,下调哺乳动物雷帕霉素靶蛋白(
mammaliantarget of rapamycin
,
mTOR
)
Ser2481
位点和丝氨酸
/
苏氨酸激酶(
AKT
)
Ser473
位点磷酸化,表明华蟾毒精抑制肿瘤新生血管形成,破坏内皮
mTOR/ HIF-1α
信号通路触发活性氧介导的血管内皮细胞凋亡
[48]
。蟾毒灵与索拉菲尼联用显著抑制肝癌
SMCC-7721
细胞裸鼠移植瘤的血管生成,明显抑制鸡胚绒毛尿囊膜和大鼠主动脉环的形成以及人脐静脉内皮细胞迁移和血管生成,下调与血管生成相关的多种细胞因子和
VEGF
,通过调节
AKT/VEGF
信号通路增强索拉菲尼的抗血管生成作用
[49]
。
2.1.5
诱导肿瘤细胞分化
大多数肿瘤细胞的分化和成熟程度都低于正常细胞,这使得它们能够快速增殖,因此,诱导肿瘤细胞分化可能是一种降低其增殖率,减少其恶性特征的良好治疗方法
[32-33]
。蟾毒灵是人骨髓白血病细胞分化的有效诱导因子,较低浓度(
0.5
~
1.0 μmol/L
)时,蟾毒灵能诱导白血病
K562
、
U937
、
ML1
和
HL60
细胞分化为单核细
胞
/
巨噬细胞样细胞,与
4'-demethylepipodophyllotoxin
ethylidene-β-
D
-glucoside
(
VP16
)、全反式视黄酸、
1α,25-
二羟维生素
D3
、重组肿瘤坏死因子
-α
联用能协同诱导白血病细胞分化
[50]
。蟾毒灵诱导人单核细胞的白血病
THP-1
细胞分化与细胞外调节激酶级联和蛋白激酶
C
同工酶相关
[51]
。
2.1.6
逆转肿瘤多药耐药
多药耐药是一种复杂的表型,多种结构上不相关的细胞毒性化合物具有耐药性,严重限制了许多常见恶性肿瘤的有效治疗,其机制与
P-
糖蛋白、多药耐药相关蛋白
1
、细胞凋亡、
DNA
异常修复和有机微环境等相关
[33]
。据报道,蟾皮中的活性成分具有逆转某些癌细胞多药耐药的能力。蟾毒灵能将
ATPbinding cassette subfamily B member 1
(
ABCB1
)过表达的结肠癌
HCT8/ADR
、
LoVo/ADR
和
HCT8/ABCB1
细胞的化学敏感性逆转至其亲本细胞水平,以剂量相关方式增加细胞内阿霉素和罗丹明
123
的积累,显著抑制
ABCB1
蛋白表达水平,并刺激
ABCB1
的
ATP
酶活性,增加阿霉素对
ABCB1
耐药的结肠癌
HCT8/ADR
细胞裸鼠移植瘤的作用,表明蟾毒灵能够有效逆转
ABCB1
介导的多药耐药
[52]
。另有研究表明蟾毒灵在
1 nmol/L
时能增强肝癌
BEL-7402/5-FU
细胞的化学敏感性,逆转倍数为
3.8
倍,显著将细胞周期阻滞于
G
0
/G
1
期,通过增加
Bax/Bcl-xL
比例诱导细胞凋亡,下调多耐药蛋白
1
以抑制药物外排泵活性,并减少胸苷酸合成酶的表达
[53]
。华蟾毒精也可显著提高
P-
糖蛋白底物药物对
P-
糖蛋白过表达结肠癌
LoVo/ADR
、
HCT116/L
和
Cao-2/ADR
细胞的敏感性,同时对其亲本细胞无影响,显著增强阿霉素对过表达
P-
糖蛋白细胞
LoVo/ADR
裸鼠移植瘤的作用,增加了化疗药物的细胞凋亡和多药耐药细胞内阿霉素和罗丹明
123
的积累,进一步机制研究发现是
P-
糖蛋白
ATP
酶的非竞争性抑制,并不改变
P-
糖蛋白的表达
[54]
。
2.2
抗炎与免疫调节作用
2.2.1
抗炎作用
在癌症相关炎症中,主要炎性基因产物的表达受核因子
κB
(
nuclear factor kappa-B
,
NF-κB
)调控。蟾皮中活性成分蟾毒灵(
0.3
、
0.6 mg/kg
)能有效抑制卡拉胶诱导的大鼠足肿胀,下调一氧化氮合成酶、环氧合酶
-2
、白细胞介素
-1β
(
interleukin-1β
,
IL-1β
)、白细胞介素
-6
(
interleukin-6
,
IL-6
)和肿瘤坏死因子
-α
(
tumornecrosis factor-α
,
TNF-α
),显著抑制
NF-κB
信号通路
[55]
;在
BALB/c
小鼠哮喘模型中,蟾毒灵可减少肺组织炎性细胞浸润和杯状细胞增生,并抑制
NF-κB
和
p-p65
蛋白表达,提示蟾毒灵可能通过抑制
NF-κB
活力发挥其抗炎作用
[56]
。此外,吲哚烷胺类生物碱(
N
-methyl serotonin
、蟾毒色胺、脱氢蟾蜍色胺和蟾蜍噻咛)也能通过抑制
TLR4/MyD88/NF-κB
和
TLR4/MyD88/MAPKs
信号通路发挥抗内毒素炎症作用
[57]
;蟾皮水提取物、蟾皮
60%
乙醇提取物和蟾毒色胺均能有效抑制脂多糖和
phorbol12-myristate 13-acetate
(
PMA
)刺激的
U937
细胞促炎细胞因子
TNF-α
和
IL-6
的分泌,这与抑制
NF-κB
的激活有关
[58]
。
Zhang
等
[24]
从蟾皮亲水成分中分离得到的总吲哚烷胺类生物碱在脂多糖刺激的斑马鱼胚胎中表现出显著的抗炎活性;质量浓度为
50
、
100 μg/mL
时,抗炎率分别为
35%
、
55%
,优于阳性药物消炎痛;其中
N
-methyl serotonin
能显著减少中性粒细胞的数量,呈剂量相关效应,质量浓度为
100 μg/mL
时,抗炎率为
60%
;而蟾蜍噻咛的抗炎活性相对较弱,质量浓度为
100 μg/mL
时,抗炎率仅
30%
。
2.2.2
免疫调节作用
哺乳动物免疫系统的主要功能是监测阻滞内稳态,防御病原体的入侵或感染,清除受损细胞等,包括癌症在内的许多疾病通常与免疫系统受抑制有关
[33]
,因此,急需增强免疫系统功能的药物。研究表明,蟾皮中的蟾蜍二烯羟酸内酯类化合物有增强免疫系统功能,蟾毒灵(
5
、
10 mg/kg
)显著减弱哮喘模型小鼠的高反应性,抑制气管肺泡灌洗液中总炎症细胞数,包括巨噬细胞、嗜酸性粒细胞、淋巴细胞和中性粒细胞,通过降低血清中
Th2
相关细胞因子(如
IL-4
、
IL-5
、
IL-13
和卵清蛋白特异的
IgE
)的释放来干扰
Th2
介导的免疫反应
[56]
。华蟾毒精促进小鼠淋巴细胞增殖,显著提高
CD4
+
/CD8
+
T
细胞比例,促进淋巴细胞从
G
0
/G
1
期向
S
期转化,为
S
期向
G
2
/M
期转化提供必要条件;增强
IL-2
和
IL-10
的分泌,并提高巨噬细胞的吞噬能力
[59]
。
Wu
等
[60]
研究显示,华蟾毒精可通过增强
Th1
免疫反应,显著增强福尔马林灭活的沙门氏菌疫苗对小鼠的保护作用;显著促进小鼠脾细胞增殖,上调干扰素
γ
(
infectionrequires interferon-gamma
,
IFN-γ
)和一氧化氮表达,并增加沙门氏菌免疫小鼠血清中沙门氏菌特异的总免疫球蛋白
G
(
immunoglobulin
,
IgG
)和
IgG2a
的表达水平;在体内,华蟾毒精能减少小鼠脾脏的菌落数,并延长沙门氏菌免疫小鼠感染活鼠伤寒沙门氏菌的存活时间。华蟾毒精还能有效抑制脂多糖诱导的树突状细胞成熟和多种细胞因子(
TNF-α
、
IL-6
、
IL-8
、
IL-10
和
IL-12p40
)产生,激活
caspase-1
并显著增强
IL-1β
的产生,上调树突状细胞中抗菌肽人
β-
防御素(
human β-defensin
,
hBD-2
)和
hBD-3
的基因表达,诱导中性粒细胞分泌人中性粒细胞多肽
1-3
(
humanneutrophil peptides 1-3
,
HNP1-3
)和抗菌肽
hCAP-18/LL-37
,以增强中性粒细胞的抗菌活性
[61]
。远华蟾毒精能增强小鼠对卵清蛋白的
Th1
免疫反应,增强
Th1
转录因子
T-bet mRNA
表达水平,影响
IFN-γ
的分泌,促进卵
清蛋白免疫的小鼠脾细胞增殖,
CD3
+
/CD4
+
、
CD3
+
/
CD8
+
升高,证实远华蟾毒精对细胞免疫应答有增强作用
[62]
。
2.3
抗菌与抗病毒
蟾皮总生物碱为蟾皮的抑菌活性成分之一,蟾皮总生物碱的含量随乙醇浓度的升高而增多。王元清等
[63]
采用滤纸片法测定了蟾皮不同溶剂提取物的抑菌性能,结果发现,
90%
乙醇提取物的抑菌效果强于
60%
乙醇提取物,
90%
乙醇提取物对金黄色葡萄球菌、大肠杆菌、枯草芽孢杆菌和黑曲霉菌的最小抑菌浓度分别为
1000
、
500
、
500
、
500 mg/mL
,且抑菌活性部位具有一定的耐热耐压稳定性。此外,远华蟾毒精和海蟾蜍精对大肠杆菌的最小抑菌浓度分别为
64.0
、
16.0 μg/mL
,对金黄色葡萄球菌的最小抑菌浓度均为
128 μg/mL
[64]
。
在乙型肝炎病毒感染期间,乙肝病毒
e
抗原(
hepatitis Be antigen
,
HBeAg
)促进免疫耐受和持续感染。
Cui
等
[65]
研究发现,超临界二氧化碳蟾皮萃取物浓度低于
0.01 μg/mL
时对
HepG2.2.15
细胞无显著细胞毒性作用,用
0.1 ng/mL
萃取物处理
HepG2.2.15
细胞
6 d
后,能有效抑制
HBeAg
和乙肝病毒核心抗原(
hepatitisB core-related antigen
,
HBcrAg
)的分泌,抑制率分别为
23.36%
、
30.94%
,均高于阳
性药物
3CT
(
20.81%
、
17.38%
)并且
HBV
mRNA
表达水平也明显受抑制。蟾毒灵给药
6 d
后,在
0.1 nmol/L
时能显著抑制
HBcrAg
分泌(抑制率
19.58%
),
效果优于阳性药物
3TC
,对
HBeAg
(
11.36%
)的抑制作用高于
HBsAg
,乙肝病毒表面抗原(
hepatitis B surfaceantigen
,
HBsAg
),乙肝病毒
mRNA
表达水平下降,但对乙肝病毒
DNA
无抑制作用;而华蟾毒精给药
3 d
后,在
1
nmol/L
时对
HBeAg
分泌的抑制效果(
8.28%
)与阳性药物
3TC
(
100 μg/mL
,
7.71%
)的作用相当,对
HBeAg
的抑制作用高于
HBsAg
,乙肝病毒
mRNA
表达水平下降,同样对
HBV DNA
无抑制作用
[66]
。
蟾毒色胺可通过烟碱乙酰胆碱受体的竞争性机制抑制狂犬病毒感染仓鼠肾成纤维
BHK-21
细胞,并呈剂量和时间相关效应,质量浓度为
3.9 mg/mL
时,感染抑制率达到
100%
[67]
。与抗菌肽
ocellatin-F1
联用,具有协同抗狂犬病病毒的作用
[68]
。在小鼠体内,
0.63 mg/d
蟾毒色胺是抗狂犬病病毒感染的有效剂量,对动物的生理和中枢神经系统不会产生显著影响;高剂量(
1.05
和
2.1 mg/d
)的蟾毒色胺耐受性良好,只造成轻微的行为影响
[69]
。
2.4
强心作用
蟾皮中的蟾蜍二烯羟酸内酯类化合物属于强心甾类化合物,具有类似于洋地黄的强心作用
[32]
。陈才法等
[70]
研究表明蟾皮脂溶性成分对心肌细胞膜上
Na
+
-K
+
ATP
酶的抑制作用显著,抑制率为
44%
;蟾皮水溶性成分对心肌细胞膜上
Ca
2
+
ATP
酶活性的抑制率超过了
50%
,对
Ca
2
+
-Mg
2
+
ATP
酶活性抑制率大于
60%
。蟾毒灵在低浓度(
0.01
~
0.5
μmol/L
)范围内能有效增强离体豚鼠的心房收缩力,并对收缩频率及节律均无明显影响;在高浓度(≥
0.7
μmol/L
)时则引起心室正常收缩以外的收缩
[71]
。陈立等
[72]
将
0.133 mg/mL
华蟾毒精静脉恒速输入到心衰家兔体内后,家兔的左室内压(
leftintraventricular pressure
,
LVP
)、左室内压最大变化速率(
LVP
±
d
p
/d
t
max
)和平均动脉压(
mean artery pressure
,
MAP
)值均明显回升,且强心效力明显强于毒毛旋花子苷
K
,除此之外,华蟾毒精的最大有效剂量、最小中毒量、最小致死量和治疗指数也明显高于毒毛旋花子苷
K
。
蟾皮化学成分的药理活性见表










