前言
首先搞科研必须要有科研的心态。细心是干任何事所必须具有的,对于科研更是至关重要,细节决定了成败;信心也是要具有的,实验失败是再平常不过的事了,失败一定不要气馁,从头分析,设计对照,逐一排除各项因素,找出问题的所在,不断地改进,相信自己最终会成功的。恒心也是对于科研至关重要的,一项课题不是一天两天完成的,需要日复一日的去做,有时候就是一个实验操作要重复的做,因此只有拥有了恒心才能持之以恒的向最后的成功迈进。最后就是绝心,绝心就是指在进行任何实验操作时要持有绝对的的态度,不可有模棱两可,差不多的态度。这些是搞科研所必须具有的素质。用一句经典的话就是:复杂的事情简单做,简单的事情认真做,认真做的事情重复做,重复做的事情创造性做!
其次要掌握一定的方法。实验之前必须进行分析,制定实验计划,分析影响因素。任何实验基本上会经历以下过程:
一、查阅文献。查阅文献时先查阅中文相关文献,然后对相关内容有所了解后查阅外文文查阅外文文献可以使用图书馆数据库、NCBI中pumbed数据库、谷歌学术搜索等。
二、 设计流程及时间表。对整个实验流程事先必须要熟悉,以便根据自己的时间进行相关的时间安排,做到不急不躁,稳中求进。尤其不要忽视试剂用具准备及样品处理的时间。
三、试剂及用具的准备及样品的处理。材料用具准备的前提是对实验的步骤充分了解,了解以后据直准备。对于分子生物学相关的实验基本都需要无菌的试剂及用具,因此需要事先进行相关的灭菌处理。对于绝大多数试验的第一步都需要样品的相关处理,然后才能进行之后的相关实验操作。样品处理尤其是对某条件的多梯度处理时,一定要保证其他条件的一致性,方可具有比较性,因此处理样品时要充分考虑如何控制其他条件的一致性。准备工作是整个实验的核心,不可忽视,确保第一步必须要走好!
四、实验设计及可能影响的因素。不同的样品、不同的条件、不同的地方因此也决定了实验的不同性。运用哲学说就是具体问题具体分析,共性中有个性。因此对于某个实验操作不可完全照搬他人的实验步骤,应根据自己的实验进行相关的优化。另外每一步实验操作的的原理必须要搞懂,这是优化实验步骤和寻找问题所在的前提。
最后不要犯一些小的错误,要养成一些科研习惯。比如标记的习惯,无论个数少还是多都要按照标记的规则进行标记,尤其配制试剂时;做笔记的习惯,无论是达到了预期的结果还是失败的结果都应做好笔记,这是以后反思的重要材料来源;还有就是正常工作的习惯,一时结果不好,不要挑灯夜做,这样更不会有什么效果,错误不断,只有休息好了才能做好实验。
本文主要详细介绍了一些常规实验操作步骤及注意事项以及部分生物信息学知识等内容,其内容都是编者所在实验室常用的步骤以及编者在实验过程中所遇问题的总结,特别适合于初学者的学习。本书参考了相关的文献及程娇娇、曹京、王桂栾、毛瑞峰等的毕业论文以及网络论坛、fermentas公司操作注意事项(http://www.fermentas.com/en/support/application)等内容,在此对他们表示感谢。特别对10级生物科学薛瀚师妹DNA重组章节及荧光定量部分的修正和总结也表示感谢。另外编者推荐几个实验技术交流的论坛:生物秀论坛,生命经纬,丁香园,生物谷等,供读者学习交流使用。
由于小编水平有限,书中错误在所难免,还望以后的师弟师妹们继续补充和修改。
最后送各实验室的同志们,科研之路并不是一帆风顺的,但我们一定要坚信:struggle and persist!
第一章 DNA提取
一、CTAB法
实验原理
CTAB是一种非离子去污剂可与DNA形成复合物,此复合物在高浓度氯化钠溶液(20.7mM)下可溶,并稳定存在,但在低浓度下(0.1—0.5mM)沉淀,而大部分蛋白质、多糖仍溶于溶液中,经离心弃上清后,CTAB核酸复合物,用75%乙醇浸泡脱掉CTAB。
实验步骤:
注:夏天取新鲜叶片时应带冰盒,将取下叶片立刻放冰上,防止过度失水,另外取材尽量取用嫩组织。
1)取约1cm2的新鲜的幼嫩叶片于研钵,向研钵中加入600µl预热的 2%的CTAB分离缓冲液和12ul的β—巯基乙醇(2%),研磨成浆,倒入1.5ml的离心管中。
1.此步主要是充分破坏细胞壁,使CTAB充分接触细胞膜。
2.此方法为小量法,样品的量要必须符合要求。
3.裂解时间尽量要短。
4.CTAB溶液在低于15℃ 时会形成沉淀析出,因此,在将其加入冰冷的植物材料之前必须预热,且离心时温度不要低于15℃。
5.转移时可用剪去尖端的枪头吸取。
(2)将离心管置于65℃水浴30min,其间每隔10min轻轻摇晃离心管,使其充分混匀
使CTAB充分破坏细胞膜,与DNA结合形成复合物溶于高盐溶液。
(3)加入等体积的酚/氯仿/异戊醇(25:24:1),颠倒摇晃5min。
(4)12000r/min离心10min,收集上清
降至室温后,再加酚/氯仿/异戊醇。4℃离心更容易去蛋白,故凡是抽提蛋白的,都可运用此条件。此后不再赘述。另外收集上清时应调小枪头(50ul即可),缓慢吸取。此后不再赘述。
(5)加等体积酚/氯仿(1:1),混匀,12000r·min-1离心10min,取上清。
(6)加等体积氯仿/异戊醇(24:1),混匀,12000r·min-1离心10min,取上清
此步氯仿是为了除去上步残余的酚。
(7)向上清中加入两倍体积预先冰冻的无水乙醇或一倍体积的异丙醇,置于-20℃冰冻30min。
降低CTAB的浓度,使DNA析出到难溶的乙醇中。加入醇类使其终浓度为70%左右沉淀效果最好。若杂质含量较多,可采用室温沉淀。异丙醇沉淀较完全,但难洗涤,适合小量提取DNA沉淀。另外沉淀时间不是很严格,DNA可以在其中长期保存,但若想减少杂质含量尽量不要过长。
(8)12000r/min离心10min,弃上清,加入75%的乙醇,悬浮沉淀,常温下静置5min,弃上清。
离心之前将液体吸出,可以减少杂质的含量。但吸的时候一定注视枪头尖部,不要把DNA吸走了。另外离心获得的沉淀呈现透明状,似果冻,若有粉末状物质则可能蛋白质未抽提干净。
(9)室温干燥,溶于20µl 无菌水中,然后加入1/10体积的RNAase(1mg/ml)37℃处理40min后-20℃保存备用
晾干之前,可短暂离心,将管壁乙醇聚集底部,后用移液枪吸去,能够最大限度的减少乙醇的存在,加速晾干。晾干的标准是无酒精味即可,不可晾的时间过长。
二、SDS法
实验原理
植物组织经液氮研磨破碎后,加入细胞提取液能进一步破坏细胞,释放核酸。其中细胞提取液中的SDS可以破坏细胞膜,使蛋白质变性,EDTA可以抑制DNA酶的活性,防止DNA被降解。之后经过酚氯仿的多次抽提经离心后可以将细胞中的蛋白质、糖类清除,然后经乙醇沉淀获得DNA。
(1)取0.5g烟草叶片,洗净,吸干表面水分,在液氮中迅速研磨成粉末状(越细越好)

(2)转移至1.5mL离心管中,加入600ml预热的细胞提取液,充分混匀,65℃水浴保温20min。
(3)加等体积酚/氯仿/异戊醇(25:24:1),混匀,12000r·min-1离心10min,取上清。
(4)加等体积酚/氯仿/异戊醇(25:24:1),混匀,12000r·min-1离心10min,取上清
(5)加等体积氯仿/异戊醇(24:1),混匀,12000r·min-1离心10min,取上清
(6)加等体积异丙醇或两倍体积的无水乙醇,混匀,-20℃放置30min以上。
(7)12000r·min-1离心10min。
(8)去上清,加70%乙醇使沉淀悬浮并洗涤2次,每次12000r·min-1离心5min
(9)去上清,再短暂离心,用移液枪将残余的乙醇吸出,沉淀于室温下晾干,加20ul无菌水溶解。然后再加入1/10体积的RNAase(1mg/ml)37℃处理40min后-20℃保存备用。
参考文献
魏群.分子生物学实验指导[M].北京:高等教育出版社,1999,69-70
第二章RNA提取
本文详细介绍RNA提取的实验前准备及详细实验步骤,对于做克隆和荧光定量的同学们有所帮助!同时要时刻警惕RNA提取也做到无菌操作的水平!
Trizol法
实验原理
Trizol内含异硫氰酸胍能迅速破碎细胞,同时使核蛋白复合体中的蛋白变性并释放出核酸;由于释放出的DNA和RNA在特定pH值下的溶解度不同,且分别位于中间相和水相,从而使DNA和RNA得到分离;取出水相后,通过有机溶剂(氯仿)抽提及异丙醇沉淀,可得到纯净RNA。
预备实验
1.DEPC简介
DEPC是一种潜在的致癌物质,主要是能生成乙酯基衍生物和乙酯类衍生物,其中尿烷是一种已知的致癌物质。DEPC有刺激性,对眼睛和气道粘膜有强刺激,在操作中应尽量在通风的条件下进行,DEPC毒性并不是很强,但吸入的毒性是最强的,使用时戴口罩,不小心占到手上注意立即冲洗。由于DEPC是一种酯类,较难溶于水,故通过加热搅拌的方式来使其溶解。
2.1%的DEPC水的制备(1L)
取1mlDEPC于999ml去离子水中,37℃在磁力搅拌器上搅拌4h以上使其溶解。此溶液即1%DEPC水。
注意:
①DEPC与水反应后会产生大量的气体,可用塑料膜将瓶口扎紧,剧烈混匀后通过观察塑料膜是否胀气来判断反应是否充分
②DEPC水的量可以根据需要配制即可,原则上确保所有的物品都能浸在DEPC水中。
③制备的1%DEPC水,应留下100ml左右并装入100ml左右的试剂瓶中,以备溶解RNA用。注意:根据试剂瓶的大小调节所留的DEPC水的量,原则是确保DEPC水能够充满整个试剂瓶,并且盛有DEPC水的试剂瓶也要放置过夜,然后高温高压灭菌使DEPC分解后才能用于溶解RNA。
④配制DEPC处理水时用棕色瓶。
3.离心管、各种大小的枪头(1000ul、200/100ul、10ul)根据步骤及提取的量计算所需的个数,然后用制好的1%DEPC浸泡比计算数目略多的过夜处理。
注意①处理方法:用镊子夹住离心管,然后浸入DEPC水中,使离心管中充满液体,然后再倒出,反复2次,最后一次再浸入DEPC水后直接浸入其中,无需倒出。而枪头则用最大量程的枪吸打DEPC水2次,最后一次吸入后不再打出,直接把枪头推至DEPC水中。
②处理完毕后,离心管一般用一培养瓶盛装,枪头用枪头盒盛装,故DEPC处理时也应包含这些器皿。
4.研钵、研棒、试剂瓶、镊子、废液瓶等用锡箔纸包好口后200℃干热灭菌4h。
5.RNA酶的来源。大量的微生物存在于空气中、水中及物体的表面,当它们死亡时,会释放大量的RNAase。对于人的皮肤、粘液、唾液等都因为直接或间接的与空气接触而携带有RNAase。
实验步骤
主要参见trizol试剂的说明书,以下步骤仅供参考。
(1)取离心管做好标记,0.2g样品液氮研磨成粉末状,加1mlTrizol,漩涡混匀,室温静置5min。
可提前将离心管放液氮凉一会;一定要将材料研磨充分,且一旦研磨完毕立刻加入Trizol混匀。这一步是能否提出RNA的关键。
(2)加200µl氯仿,上下颠倒15s,室温静置3min。
提前打开冷冻离心机,若在夏季进行实验需开启制冷开关。
(3)12000 r/min 4℃离心15min。
(4)取离心管,样品分三层(无色上清水相,中间白色层,粉色下层有机层)小心吸取无色上清至另一新离心管。收集上清时应调小枪头(50ul即可),缓慢吸取。
(5)加入等体积异丙醇,轻轻混匀,冰浴10-20min。
这段时间可以用灭菌处理的DEPC水配制75%乙醇。
(6)弃上清,用75%乙醇1ml悬浮沉淀。
(7)12000 r/min 4℃离心10min。
离心之前一定要悬浮沉淀,即指用移液枪将沉淀打起。
(8)弃上清,再短暂离心,用移液枪将残余的乙醇吸出,沉淀于室温下晾干。
晾干的标准是无酒精味即可。
(9)加入30-50µlDEPC处理的水溶解RNA。
提取完毕后分出2管,其中一管装2ulRNA,用于浓度的测定;另一管装2—5ul,用于电泳检测。浓度测完后根据反转录的要求分装小管,保证每一管符合反转录反应所需的RNA的质量。若用于半定量,则必须保证各样品每管的质量均相同。
(10)电泳时用3%H2O2对电泳整个装置消毒。
(11)取1—2ul进行电泳检测及浓度检测。
(12)分装,-80℃保存。
注意事项
1. 氯仿、异丙醇、乙醇都应用未开封的,75%的乙醇用移液枪配制,勿用量筒等中间器皿。
2. 整个过程要及时更换手套,戴双层口罩,并在操作区开启酒精灯。
3. RNA一定要贮存到-70℃冰箱,在-20℃保存时间很短。
4. 配制的溶液应用灭菌处理的DEPC水。
5. 移液器: RNase的又一污染源是移液器。根据移液器制造商的要求对移液器进行处理。一般情况下采用用DEPC配制的70%乙醇擦洗移液器的内部和外部,基本达到要求。
RNA提取,对于整个分子生物学实验来说,算是比较有难度的操作了,提取出RNA如何判断质量的好坏呢?那当然通过电泳图了。本次小编将讲述电泳图上的秘密,帮助您更好的来提取RNA,提高成功概率。
结果分析
1. RNA条带不亮或缺失
(1)上样量不足。
(2)染色剂不足或染色剂分布不均匀。
(3)RNA跑出凝胶。电泳至溴酚蓝至凝胶的2/3即可停止电泳。另外电泳槽应水平放置,缓冲液要浸过凝胶,
(4)RNA在凝胶中发生扩散。避免长时间的电泳,否则小片段容易扩散。
(5)RNA降解。最大限度减少在紫外线下暴露的时间,使用新鲜的电泳缓冲液,新制的凝胶,电泳槽及制胶的模具梳子等用双氧水浸泡,高电压短时间电泳(20min)。也可能使用的组织材料不够新鲜(冷冻的材料必须在化冻之前将其磨碎)。
2. RNA条带弥散
(1)RNA被核酸酶降解。要用新的缓冲液,制胶用的模具要清洁,枪头要灭菌。
(2)电泳条件不适合。避免长时间电泳,缓冲液要浸过凝胶,电压要合适,不能过低。电压过高,造成电流过大,也容易引起此现象的发生。一般5—8V/cm.(聚丙烯酰胺电泳多采用8V/cm)所以计算采用电压时应先测量电泳槽两电极之间的距离。为了增加条带的清晰度,电泳开始的几分钟建议使用低电压。
(3)上样量过大。根据沉淀量的多少确定点样的量,一般1—3ul。
(4)胶孔不规则。插梳子时应垂直插入,凝固后拔出梳子。
3. 背景很高
(1)核酸染剂加入的量过多。应按说明书添加。
4. 点样孔发亮
(1)上样量过大。
(2)胶孔未凝固好,使样品无法往前移动。
(3)RNA样品中含有污染物,如残留的蛋白质容易引起此现象。可能trizol量太少,应加大trizol用量。另外若残留DNA也应考虑加大trizol用量。
5. 出现多条带
(1)RNA降解。提取过程中出现降解或电泳过程中出现降解(可能性较小)。
(2)上样量过大。RNA由多种类型组成,若上样量过大则会导致各种类型的RNA(tRNA,mRNA等)均会出现条带。
提取常见问题
1.研磨必须要充分,样品用量要严格。
2.处理的枪头离心管要量少多份,无RNAase的DEPC水也要量少多份,以备分装RNA和调浓度使用。
3.电泳时间10-20min,不宜过长,电压可为120-130V。
4.点样用的10ul枪头也要提前处理。
5.枪头离心管90℃烘干5——6h。
6.电泳时,可以点样Marker作阳性对照。
7.从-70℃冰箱中取出材料要在解冻之前迅速研磨。
8.提出的RNA可在70%酒精-70℃保存一年。
9.所有的东西必须有多余,防止出现用品的损失。
10.试剂瓶可以连续使用。
附:核酸提取注意事项
提取一种核酸时,首先应该查阅文献,了解是否有前人尝试此种物种核酸的提取方法,若有则可以参照其方法进行提取,否则应首先了解此种植物含糖类、酚类、次生代谢物等物质的多少,然后制定相应提取方案,以实现最大程度的去除这些物质。提取核酸材料选择是至关重要的。新鲜幼嫩的组织往往糖类、酚类、次生代谢物等杂质含量少,因此提取的核酸质量(纯度)会较高,对后续的PCR、酶切等反应有利,而老的组织由于各种杂质含量都较高,因而不易获得纯净的核酸,影响后续的各种酶促反应。
关于破碎细胞。破碎细胞一般是采用研磨法。在研磨之前将材料剪成小块,可以直接研磨也可以用液氮研磨(但提RNA必须用液氮研磨),将组织磨成匀浆即可。在这里对冷冻的材料特别要求从冰箱中取出后要迅速称量,然后研磨,在提RNA时要保证材料在液氮研磨之前不能融化。
关于称取样品质量。根据提取量的要求,选择一次研磨的质量,然后平均分至各管中(后面的步骤均为提一管所需的各试剂的量)。
酚/氯仿/异戊醇在抽提过程中的作用。酚对蛋白质的变性作用远大于氯仿,但是酚的比重略比水重,碰到高浓度的盐溶液,离心后酚相会跑到上层,不利于含质粒的水相的回收;但加入氯仿后可以增加比重,使得酚/氯仿始终在下层,方便水相的回收;另外酚与水有很大的互溶性,如果单独用酚抽提后仍有大量的酚溶解到水相中,其会抑制后续的酶促反应(比如限制性酶切反应),因此单独用酚抽提后一定要用氯仿抽提一次将水相中的酚去除,而用酚/氯仿的混合液进行抽提,跑到水相中的酚则少得多,但最好再用氯仿抽提一次,将酚完全除去。异戊醇,其作用主要是为了让离心后上下层的界面更加清晰,方便水相的回收。另外在一次酚/氯仿抽提之后,上清液仍然呈现较深的颜色,则可以考虑再次进行酚/氯仿抽提。因为在植物中存在各种色素,主要包括脂溶性色素与水溶性色素两类。脂溶性色素多为四萜类衍生物,主要有叶绿素、叶黄素、胡萝卜素、番红花素和辣椒红素等,它们易溶于乙醇,酚和氯仿等溶剂。其中胡萝卜素不溶于乙醇。水溶性色素主要为花色甙类,又称花青素,普遍存在于花中。可溶于水与乙醇,不溶于乙醚与氯仿等有机溶剂。因此一般可以通过再次酚/氯仿抽提将色素除去。
对于沉淀核酸方法的讨论。沉淀核酸主要通过有机溶剂法沉淀,原理主要是降低了介电常数,破坏了了核酸分子表面的水化层。另外还可以利用等电点法和盐析法沉淀,如利用醋酸钠(3M,pH5.2),一方面高盐浓度破坏核酸分子水化层另一方面使其接近核酸等电点。(核酸的等电点比较低。如DNA的等电点为4~4.5,RNA的等电点为2~2.5)。常用的有机溶剂是乙醇和异丙醇,它们的区别是:沉淀DNA 通常使用冰乙醇,在低温条件下放置时间稍长即可使DNA沉淀完全。而沉淀RNA通常使用异丙醇。异丙醇沉淀速度快但通常把盐分也沉淀下来。在使用量上,乙醇终浓度为70%即之前的溶液中加入2—2.5倍体积的无水乙醇,使其浓度达到70%左右,而异丙醇一般使用等体积的量。之后的70%乙醇起到洗涤和再次沉淀的作用。
关于沉淀结束后的操作。沉淀结束后,若沉淀量较大,可以考虑用枪头或灭菌牙签等将沉淀“钩出”,放入新的离心管中,然后用70%乙醇洗涤;若沉淀量较少,沉淀结束后可以将无水乙醇最大量的吸出然后离心。这样可以明显减少提取核酸中盐分、糖类等杂质的含量。
关于乙醇洗涤。常用70%浓度可以有效去除盐离子另外可以洗涤两次,效果会更好。洗涤完成后,必须要晾干,才能进行溶解,防止残留乙醇影响后续的酶促反应。
关于溶解核酸溶剂的选择。当想长期保存,暂时不进行下一步实验时,采用TE缓冲液,其中的Tris-HCl的提供一种缓冲系统,EDTA可以螯合二价金属离子进而抑制依赖于二价金属离子的DNA酶的活性,对DNA起到保护作用。
第三章 PCR
整天谈论测序,那测序里面关键的一环是什么呢?当然是PCR了。呵呵,本次小编就写给只会做分析而不会做实验的的生物信息小伙伴们。很重要奥,所有的分析最终都要落到验证上奥!
一、实验原理
PCR是体外人工选择性扩增DNA的一种方法,它类似于生物体内DNA的复制。在生物体内DNA复制需要模板、引物、DNA聚合酶、DNA解旋酶、 dNTPs;而体外PCR反应同样需要类似的成分,有模板、引物、PCR Buffer、Taq酶、dNTPs。其中引物是人工设计的特定序列,实现对特定位置的扩增;PCR Buffer提供一个反应的缓冲环境;反应过程同生物体内一样,DNA双链打开,引物结合模板,延伸形成新链。而这些过程在生物体内靠DNA解旋酶解链,而在体外在通过控制反应温度实现的。如常用94℃变性模板DNA打开双链,在退火温度处引物结合到模板上,最后在72℃完成延伸,并反复重复此过程即可实现对特定片段DNA的大量扩增。到第三循环开始才产生出和靶DNA区段相同的DNA分子,进一步循环地产生出靶DNA区段的指数加倍。
在扩增后期,由于产物积累,使原来呈指数扩增的反应变成平坦的曲线,产物不再随循环数而明显上升,这称为平台效应。平台期(Plateau)会使原先由于错配而产生的低浓度非特异性产物继续大量扩增,达到较高水平。因此,应适当调节循环次数,在平台期前结束反应,减少非特异性产物。到达平台期(Plateau)所需循环次数取决于样品中模板的拷贝。
二、主要的成分
模板
1.模板可以是多种形式,主要包括基因组DNA、质粒DNA、携带病毒的基因组DNA、PCR产物,cDNA等,但不能为RNA。对于不同类型的模板,其主要不同在于预变性的时间以及模板的量。一般对于大的基因组DNA预变性时间10min足够,质粒DNA2min、携带病毒的基因组DNA预变性2min、PCR产物预变性2min即可。
注意cDNA为单链DNA,但仍可做PCR的模板,只不过在第一轮循环时只有一个引物结合,合成另一条链。从第二轮开始,两个引物均与特异位点结合,从而实现了与常规PCR的接轨。而同作为单链的RNA却不能进行PCR扩增,原因就在于实现PCR反应的是DNA聚合酶,只能特意识别DNA链。
2.对于模版的量来说,一般25ul体系DNA的质量为50—100ng。对于基因组DNA由于其结构比较复杂,提取的浓度也往往比较大,为防止浓度过大对PCR造成影响,故需对提取的DNA进行梯度稀释。否则浓度过高则可能会引起非特异性扩增。对于质粒及PCR产物由于其结构简单,且提取浓度一般较低,则无需稀释。
引物
1.一般引物用贮液浓度为10uM。对于刚合成的引物可以根据期管壁上的说明计算稀释至10uM即可(一般引物说明上配制的浓度往往过高,所以必须稀释至10uM才能利用实验室最小刻度(0.5ul)的移液器取到)。另外引物一旦稀释完毕,应注意分装小份,防止在用的过程中反复冻融引物而造成引物的失效。
2.一般25ul反应中引物用量为0.5ul(终浓度要求为为0.1——1uM),若引物若放置时间比较长,则可以适当增加引物的用量。引物用量过多,容易导致引物二聚体(电泳时位于前方不成条带的小片段)的出现。
附:
1.引物稀释的方法:配制时首先将合成的引物12000r/min离心5min,室温静置1分钟,然后再慢慢打开管盖,根据引物合成单(或储存引物的离心管)说明加入一定量的灭菌水(引物说明是加入Buffer,此处加入无菌水即可)以达到10uM的浓度,在漩涡振荡器上充分振荡5-10min,即配成10um的贮存液,-20℃保存备用
PCRbuffer
一般PCR Buffer为10×的,使用时终浓度应为1×。如25ul体系中,应加入10×PCR Buffer为2.5ul。另外使用时应注意是否添加了Mg2+,若没有,则需要再单独添加,有则无需。
Mg2+
推荐用量为1—4mM。浓度过低,影响PCR产物的产量,而浓度过高,则出现非特性扩增。在PCR反应中Mg2+浓度,需进行优化。
三磷酸脱氧核苷酸(dNTPs)
避免反复冻融,应分装小份。dNTP的浓度取决于PCR反应体系中的特定靶序列的长度,常用浓度为0.2 mM。据此我们可以确定反应体系中的dNTP的最低适宜浓度。当dNTP的浓度大于50mmol/L时会抑制Taq酶的活性。需要注意的是dNTP与Mg2+之间的浓度关系。因为dNTP中的磷酸基团可与Mg2+结合,使反应体系中游离Mg2+浓度下降从而影响Taq 酶的活性。
Taq酶
PCR使用的Taq酶在反应时由于丧失3’——>5’外切核酸酶活性,因而不具有校正活性。在典型的一次PCR反应中,taq酶造成的错误的核苷酸错误的掺入率大概为每20000个核苷酸中就有一个,虽然表面上看起来不会有多大的影响,但是在分子克隆中如果有一个错误核苷酸掺入,很有可能造成移码突变,影响是严重的。为弥补taq酶这一缺点常将克隆后的序列进行测序,来确认所扩增序列的准确性。同时也可采用保真度更高的Pfu DNAPolymerase,来确保扩增的准确性,Pfu DNAPolymerase有3 ’-5’的外切酶活性,5’-3’外切酶活性,是目前已发现的所有耐高温DNA 聚合酶中出错率最低的。但PCR产物为平端,不能进行Ta克隆!而解决此问题的办法是使用TaqPlus DNAPolymerase,该酶是Taq 酶和pfu Polymerase的混合物,因此既能保证准确性又能保证能够进行Ta克隆。在使用中应该注意如下问题:
1.25ul体系中一般用量为0.1ul(用枪头沾一下即可),用量过多可能会出现非特异性扩增。
2.PCR体系构建过程中最后加入的一种成分,因此只有在其他试剂加完之后才可从冰箱中取出加入。
3.Taq酶中含有甘油,故不结冰,若结冰则应丢弃。
4.对于预变性时间较长的PCR反应,应在预变性即将结束后加入酶,减少长时间高温对酶活力造成的损伤。
5.保存时间过长的酶可以适当增加用量。
三、体系构建
1.加样原则是酶最后加,倒数第二加的为模板。加样时应从离心管底部将各成分依螺旋式上升的方式加到管壁上。如果做多管PCR,那么按照如下方法计算出公共成分做n管时所需总体积然后混合均匀之后再分装到各个离心管中去,这样可以有效地避免误差及污染。
V总 =V标×(n+1)
其中V总需配制的总体积V标每管PCR反应需要某成分的标准体积,n表示所做PCR的管数。
2设立适当的阳性对照和阴性对照,检测PCR各试剂的可用性及污染性,便于对电泳结果准确的分析。
四、预实验
准备一个冰盒,并将做PCR的各试剂(除酶)、模板及无菌水放到冰上融化,设置PCR仪上反应程序。
五、实验步骤
下列步骤均在冰上操作
1. 取1.5ml离心管,加入各公共成分(除酶)的V总,然后分装于各PCR管中,最后将酶加入。
2. 瞬时离心10s,将各成分混匀。
3. 放入PCR仪反应。
4. 4℃冰箱保存。
5. 电泳检测。
注:常用于PCR的DNA聚合酶多要求反应必须冷启动,即反应必须在瞬间从低温达到变性温度(一般94℃)。但有些酶需要热启动,则无需在冰上操作,但酶同样还是需要现用现取,保持酶的最大活性。
示例 PCR反应体系(25µl)及条件如下:
质粒DNA 0.5µl
上游特异性PCR引物 0.5µl
下游特异性PCR引物 0.5µl
10×PCRbuffer 2.5µl
Mg2+ 1.5µl
dNTP 0.5µl
Taq PCR聚合酶 0.1µl
灭菌水 18.9µl
预变性 94℃ 2min
变性 94℃ 30s
退火 59℃ 30s
延伸 72℃ 1min
循环数 30 cycles
后延伸 72℃ 10min
附:PCR反应参数
1.预变性。预变性的目的就是破坏DNA的二级结构,使DNA充分变性,以便于引物与模版的结合,尤其对于结构复杂的基因组DNA,这一步更是必不可少的。
2.循环阶段。
①变性:一般94℃,具体参照酶说明书。
②退火:引物与模版结合。其中退火温度会由于引物的不同而有所不同的。退火温度的计算可以有两种方法。一是引物设计时引物设计软件提供了一个参考的退火温度可以参考使用。二是Tm 值也可以根据 Tm=4(G+C)+2(A+T)计算。两种引物的Tm值往往在引物合成单上被注明,故可以直接查阅。在确定退火温度时,往往是使用Tm低的引物的Tm减去5℃作为退火温度的一个基点向上向下(±10°C)摸索退火温度。为提高特异性,也可在最初的5个循环使用Tm高的引物作为Tm,后面的循环使用Tm低的。由于低的退火温度可以使引物与模板很好的结合,高的退火温度可以提高特异性,故在确定退火温度时应设置梯度,并不断优化,以确定最终的退火温度。对于丰度极低的模板的PCR,特别是稀有RNA模板的反转录PCR(RT PCR)得到的cDNA模板,在最初的1~5个循环可以用更低的退火温度,然后再行正常的退火温度,这样能明显提高PCR的成功率。退火温度高为什么特异性好呢?因为退火温度高非特异性结合会使引物与模板结合不牢固而容易解离下来,不能进行扩增,因而只有特异结合引物扩增模板,获的目的条带
③延伸:一般72℃,对于扩增长片段(>=3kb)的PCR,一般采用68℃。延伸速度一般为1kb/min,具体的参见酶说明书。
④关于循环次数。一般采用25—35次,不可采用过多的循环次数,达到需要的量即可。具体要参照模板的量和希望得到的PCR产物的产量。
3.后延伸。循环结束之后最后的一步延伸主要起的作用是:
①使PCR反应完全,从而提高PCR产量。
②产物末端加A尾的作用,因而可以直接用于TA克隆。
第四章电泳
一、电泳技术原理(以琼脂糖凝胶电泳为例)
1、首先介绍使用的载体--琼脂糖。
琼脂糖是线性的多聚物,基本结构是1,3连结的β-D-半乳呋喃糖和1,4连结的3,6-脱水α-L-半乳呋喃糖。琼脂果胶是由许多更小的分子组成的异质混合物。琼脂糖在水中一般加热到90℃以上溶解,温度下降到35-40℃时形成良好的半固体状的凝胶,这是它具有多种用途的主要特征和基础。琼脂糖凝胶性能通常用凝胶强度表示。强度越高,凝胶性能越好。质量较好的琼脂糖强度通常在1200克/cm2以上(1%胶浓度)。
为啥用它呢,因为他有很多优点奥!
琼脂糖的凝胶性是由存在的氢键所致,凡是能破坏氢键的因素都能导致凝胶性的破坏。琼脂糖具有亲水性,并几乎完全不存在带电基团,对敏感的生物大分子极少引起变性和吸附,是理想的惰性载体。天然琼脂(agar)是一种多聚糖,主要由琼脂糖(agarose,约占80%)及琼脂胶(agaropectin)组成。琼脂糖是由半乳糖及其衍生物构成的中性物质,不带电荷,而琼脂胶是一种含硫酸根和羧基的强酸性多糖,由于这些基团带有电荷,在电场作用下能产生较强的电渗现象,加之硫酸根可与某些蛋白质作用而影响电泳速度及分离效果。在琼脂糖制备过程中需要把琼脂果胶尽量去除,否则琼脂糖有可能存在极微量硫酸根和丙酮酸取代电离基团,就会造成电内渗(EEO),电内渗对质点的移动产生影响,因而质量较好的琼脂糖硫酸根含量比较低,通常在0.2%以下,电内渗比较小,通常在0.13以下。这也就是琼脂糖比琼脂贵那么多的原因了。此外,琼脂糖凝胶还有以下特点:
(1)琼脂糖凝胶电泳操作简单,电泳速度快,样品不需事先处理就可以进行电泳。
(2)琼脂糖凝胶结构均匀,含水量大(约占98%~99%),近似自由电泳,样品扩散较自由,电流对样品吸附极微,因此电泳图谱清晰,分辨率高,重复性好。
(3)琼脂糖透明无紫外吸收,电泳过程和结果可直接用紫外光灯检测及定量测定。
(4)电泳后区带易染色,样品极易洗脱,便于定量测定。制成干膜可长期保存。
总之琼脂糖凝胶电泳常用于分离、鉴定核酸,如DNA鉴定,DNA限制性内切核酸酶图谱制作等。由于这种方法操作方便,设备简单,需样品量少,分辨能力高,已成为基因工程研究中常用实验方法之一。
2、琼脂糖凝胶电泳原理
DNA分子在常规的电泳缓冲液(高于等电点的pH溶液)中5’磷酸的氢离子解离被中和,而只剩磷酸根离子,故带负电,因而在电场中向正极移动(电荷效应)。又由于琼脂糖凝胶具有分子筛效应,在一定的电场强度下,DNA分子的迁移速度取决于分子本身的大小和构型。因此琼脂糖凝胶电泳不仅可分离不同分子量的DNA,也可以分离分子量相同、但构型不同的DNA分子,如质粒三种构型。
在构型相同条件下,具有不同分子量(碱基对)的DNA片段迁移速度不一样,DNA片段迁移距离与分子量(碱基对)的对数成反比,所以可以通过已知大小的标准物移动的距离与未知线性DNA片段的移动距离进行比较,便可测出未知DNA片段的大小。同时由于电荷效应存在,因此在测定DNA分子大小时,可以提高凝胶的浓度和降低电压,减少电荷效应,增强分子筛效应而提高测量准确性及电泳的分辨率。
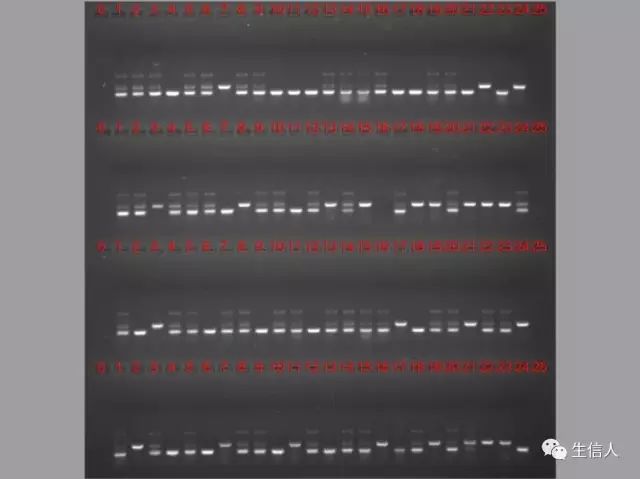
3、明白了上述原理那看图吧!
(1)线性染色体DNA
其广泛分布于动植物细胞中,为染色体的主要成分,多呈现双螺形。其电泳结果多呈现一条带,接近点样孔。若未用RNAase处理,则最前方很亮的为RNA。
(2)质粒DNA
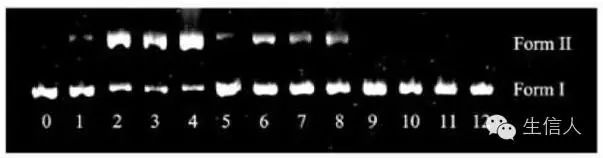
一般提取的质粒有3种构型:
1、超螺旋的共价闭合环状DNA(covalently closed circular DNA,简称cccDNA),
2、开环DNA,即共价闭合环状质粒DNA有一条链断裂,(open circular DNA,简称ocDNA),
3、线状质粒DNA,即质粒DNA 在同一处两条链发生断裂(linear DNA,简称LDNA)。
由于这3种构型的质粒DNA分子在凝胶电泳中的迁移率不同,其中超螺旋质粒DNA泳动最快,其次为线状DNA,最慢的为开环质粒DNA。通常提取的质粒DNA呈现两种构型,即电泳后呈现两条或一条带。
(3)总RNA
提取的总RNA一般呈现三条带,由于rRNA在细胞中含量最高,且提取的rRNA的质量可以反映总RNA提取的质量,故可以根据rRNA(真核生物)的三种形式5S,18S,28S的质量进行判断。如果28S的亮度在18S条带的两倍以上,我们认为RNA的质量是好。
注:
S为沉降系数(sedimentationcoefficient),当用超速离心测定一个粒子的沉淀速度时,此速度与粒子的大小直径成比例。5S含有120个核苷酸,16S含有1540个核苷酸,而23S含有2900个核苷酸。而真核生物有4种rRNA,它们分子大小分别是5S、5.8S、18S和28S,分别具有大约120、160、1900和4700个核苷酸。rRNA是单链,它包含不等量的A与U、G与C,但是有广泛的双链区域。在双链区,碱基因氢键相连,表现为发夹式螺旋。
二、设备与试剂。

1、电泳装置主要包括两个部分:电源和电泳槽。
电源提供直流电,在电泳槽中产生电场,驱动带电分子的迁移。
电泳槽可以分为水平式和垂直式两类。
垂直板式电泳是较为常见的一种,常用于聚丙烯酰胺凝胶电泳中蛋白质的分离。电泳槽中间是夹在一起的两块玻璃板,玻璃板两边由塑料条隔开,在玻璃平板中间制备电泳凝胶,凝胶的大小通常是12cm 14 cm,厚度为1mm~2? mm,近年来新研制的电泳槽,胶面更小、更薄,以节省试剂和缩短电泳时间。
2、试剂。 DNA要游泳,则必须要有水,用的水那是什么呢?比较常用就是这两种了:TAE(Tris/Acetic/EDTA)和TBE(Tris/Borate/EDTA)
1)TAE
配制:一般配制50×的母液,使用时将其稀释至1×。
特点:TAE是使用最广泛的缓冲系统。其特点是超螺旋在其中电泳时更符合实际相对分子质量(TBE中电泳时测出的相对分子质量会大于实际分子质量),且双链线状DNA在其中的迁移率较其他缓冲液快约10%,电泳大于13kb的片段时用TAE缓冲液将取得更好的分离效果,此外,回收DNA片段时也易用TAE缓冲系统进行电泳。TAE的缺点是缓冲容量小,长时间电泳(如过夜)不可选用,除非有循环装置使两极的缓冲液得到交换。
应用:大片段核酸分子分离,琼脂糖凝胶电泳DNA的回收,酶切检测,PCR反应。
2)TBE
配制:常配制成5×的母液,使用时稀释至0.5×使用(配制成5×的可以放置较长时间而不形成沉淀)。
特点:高浓度储存液易发生沉淀,故母液的配制量一次不宜过多,一旦配出后应尽快用完。
应用:对于一般的仅做检测的电泳常用此缓冲液,如DNA、质粒、RNA提取后的检测。
3)TAE与TBE的区别
TAE是Tris-乙酸,缓冲容量小,但是溶解度大,易于储存。采用TAE长时间电泳发热大,迁移速度较快,利于长片段的电泳。
TBE是Tris-硼酸,缓冲容量大,但是溶解度小,不易长期储存,易产生沉淀。其对小于1kb的片段分离效果较好,但因与琼脂糖相互作用生成非共价结合的四羟基硼酸盐复合物而使DNA片段的回收率降低,而且硼酸盐 离子有可能影响后续的酶促反应,因此回收电泳一般不推荐TBE电泳。
4)上样缓冲液(Loading Buffer)。
DNA分子在电泳中如何让他停下来,不让它从我们的凝胶中跑出去,这时就要实时监控了!除了实时监控,还必须让他沉下去,才能泳动!哈哈,用啥呢?常用的有6×的和10×的loading buffer了,一般在购买的酶制剂中均带有。下面小编就详细介绍了!
作用:第一,上样缓冲液具有指示剂溴酚蓝和二甲苯酚,它们起到指示的作用,显示电泳的进程,以便适时终止电泳;第二,里边的成分甘油可以加大样品密度,使样品密度大于TAE,从而沉降到点样孔中,防止样品飘出点样孔。另外,10×Loading Buffer是加有SDS的。SDS主要是促使聚合酶变性,因为没有除尽的聚合酶会结合在DNA双链上影响它的迁移速率。
使用上:两种缓冲液均可用于DNA和RNA电泳,但更有所侧重。6×LoadingBuffer,向电泳样品溶液中加入1/6量即可进行凝胶电泳,常用于DNA电泳。10×Loading Buffer是用于琼脂糖凝胶电泳的10倍浓度的核酸样品用,常用于RNA凝胶电泳。由于该制品中含有SDS, 容易起沫,故点样时应注意。此外其可以用来停止各种酶促反应。向酶促反应液中加入1/10体积的本制品,即可停止反应。
指示剂:10 × loading buffer 中仅有色素溴酚蓝(Bromophenol Blue)一种染料(浓度0.05%);6 X loading buffer 中含色素溴酚蓝(Bromophenol Blue)、二甲苯腈蓝FF(Xylene Cyanol FF)两种染料(浓度都是0.05%),溴酚蓝在琼脂糖凝胶(1%)中约与300 bp的双链线状DNA的迁移速度相同,而二甲苯腈蓝FF则与4000bp的双链线状DNA的迁移速度相等(不同浓度凝胶指示剂位置不同)
三、具体步骤
1、1%琼脂糖凝胶电泳胶液的制备:称取0.2g琼脂糖,置于三角瓶中,加入20ml 1×TAE缓冲液,置于微波炉中30s+10s+10s加热至琼脂溶解
加热溶解胶时待胶中起泡即可。
2、胶板的制备:将有机玻璃内槽洗净,晾干,放入制胶模具中,向冷却至65℃(不烫手)左右的琼脂糖凝胶中加入1ul Goldview后倒入有机玻璃内并在固定位置插入梳子。Goldview可以不按说明书要求加,如20ml琼脂糖加0.5ul就可以了,尤其用于回收的胶,减少goldview的量可以减少对DNA的损伤。
3、应在室温下慢慢凝固,形成均匀的孔洞,若在冰上使其迅速凝固,形成的孔洞不均匀,影响电泳
4、表面形成均匀的胶层,室温静置30min左右,凝固完全后,轻轻拔出梳子,将胶放入装有缓冲液的电泳槽中
5、加样:取5ul基因组DNA与1.2ul6×Loading Buffer 混匀后用移液枪点样,每加完一个样,换一个加样枪头,最后再点5ul的Marker 2000
1)加样时用两只手操作,以防晃动。
2)若枪头前有气泡,应小心的将样品推至枪头的最尖端。
3)注意枪头的尖端不可过深的插入孔道,以免将胶孔刺破。
4)DNA marker,每一条带都是具有固定浓度(参见说明书),电泳完毕后,就可将目的DNA条带和MARKER 条带的亮度做个大致的比较,找出两者最接近亮度的条带。在通过目测DNA浓度的时候,最好要把加样量设一个梯度,比如从0。5、2、4、6等,另外此方法也是判断紫外分光光度仪测量核酸浓度准确性的一个依据。
6、电泳:一般在80-100v电压下电泳,电泳直至溴酚蓝指示剂到胶的三分之二处停止电泳。
7、观察、拍照:将其放在紫外反射投射仪中观察,并在凝胶成像仪中拍摄电泳图片。
四、注意事项。
1、胶中所加缓冲液应与电泳槽中的相一致,溶解的凝胶应及时倒入板 中,避免倒入前凝固结块。倒入板中的凝胶应避免出现气泡,以免影响电泳结果(若出现气泡,可用带枪头的枪将气泡戳破)。
2 、 有时需要大的上样孔, 可用胶带把几个相邻的梳齿粘在一起, 这种情况下, 尽量让胶液冷一些再倒胶。
3、如果只是为了看结果,不需要拍照或回收, 同一块胶可重复用1-3次.
4、用保鲜膜包裹凝胶后放入4 度冰箱,或将凝胶浸泡在缓冲液中可保存数天。
5、若点样后胶空中的样品迅速消失,则证明胶孔具有漏洞,原因可能凝固时间太短(凝固时间至少30min)。
6、电泳缓冲液可以重复利用但必须能够保持电泳所需的一定的离子强度和pH,否则应及时更新电泳缓冲液(尤其在恒压条件下,电流过大时)。
7、为了防止电泳时两极缓冲液槽内pH和离子强度的改变,可在每次电泳后合并两极槽内的缓冲液,混匀后再用。
8、保证缓冲液淹过胶的上端1-2mm。缓冲液过多,电流则会从胶上部通过而不会通过凝胶,这样会增加分离核酸所需的时间。
9、薄的凝胶电泳结果要好于厚凝胶,但易碎应小心操作
10、一般配1×TAE时不够严格时,如浓度过大,电流就会增大.如果配得较稀,则电流会减小.另外电压与电流的比例还会受电泳槽的长宽比的影响,如果电泳槽宽固定,长度增加,则电流与电压的比会降低.
11、注意电泳期间,电泳槽盖要安全盖好,以防止液体蒸发,又可以降低被电击的可能性。
12、电泳至溴酚蓝至凝胶的三分之二时,即可停止电泳(实际过程中,跑至二分之一略多一点即可停止电泳)。
五、结果分析
1、DNA电泳的MARKER为什么是扭曲的? 电泳时电压过高,可以在电泳前15分钟用较低电压(3V/cm),等条带出孔后比较漂亮了.然后再调电压。另外上样时等样品自然沉降后再加电压。
2、电泳中产生的热通常是由中心向外周散发的,所以介质中心温度一般要高于外周,尤其是管状电泳,由此引起中央部分介质相对于外周部分粘度下降,摩擦系数减小,电泳迁移速度增大,由于中央部分的电泳速度比边缘快,所以电泳分离带通常呈弓型。降低电流强度,可以减小生热,但会延长电泳时间,引起待分离生物大分子扩散的增加而影响分离效果。所以电泳实验中要选择适当的电场强度,同时可以适当冷却降低温度以获得较好的分离效果。
3、小片段的DNA电泳应采用聚丙烯酰胺凝胶电泳以提高分辨率,上样量过多会造成加样孔超载,从而导致拖尾和弥散,对于较大的DNA此现象更明显。
4、凝胶未完全凝固,点样孔未固定好,导致条带不规则。
5、加样孔发亮:其中最主要的是非蛋白质/非核酸类的大分子杂质 (如多糖、多酚) 的残留以及蛋白质和核酸的同步残留。另外,样品使用过量,也是一个重要原因。
6、胶的厚度增大会导致电压过大
六、电泳胶图自动编号软件
1. 前言
笔者一直是做数据分析的,基本没跑过胶,前两天为了实验需要,做了几板胶图。但是胶图读带数胶孔的时候,我有些强迫症,每次数到一半就数乱了还要再重新数,一个胶图要重数好几次,这让我抓狂了。于是我决心写个程序来自动把编号添加到胶图上,方便读带。经过两天的努力写了一个自动加编号的程序,结果如下图,效果还是很好,数字位置标记很准确,再也不用担心胶图读带了。
虽然说自动,但是需要用鼠标点几下,是半自动的。我也想过通过模式识别,识别出胶孔,但是很难,而且胶图的情况复杂,全自动不现实。
程序是用R语言写的,需要使用者稍微有点R语言基础。本文就不介绍R语言程序的安装了。
2. 安装
使用前先将add_num文件夹下所有内容下载到本地(https://pan.baidu.com/s/1dFolTZj)。
该程序运行前需要安装一些R包,所需要的R包已经放在add_num文件夹下的‘1_installPackage’文件夹里。
打开R语言界面,将‘1_install.R’的代码复制到R命令窗口运行,会自动弹出一个文件选择对话框,选择‘1_installPackage’文件夹下任意一个文件即可,就会自动安装所需的包(如图)。
当出现“Congratulates”的字样时(如下图),表明R包安装成功,可以进行下一步了。R包只需要安装一次即可。
3. 使用
代码主要在2_add_num.R 里,有个 add_num()的函数。
用法:
add_num(infile,nrow=1,start=0,end=25,cex=2)
参数:
infile: 胶图文件名,支持png,tiff,jpg格式。
nrow: 添加编号的行数, 默认1。
start: 起始编号,默认0。
end: 最大编号,默认25.
cex: 字体大小,默认2.
下面讲主要的步骤了!
1、运行 source(file.choose())
在弹出的对话框中选择2_add_num.R
2、运行下面代码
add_num(infile=file.choose(),nrow=4,cex=1.5)
在弹出的对话框中选择需要添加编号的胶图(胶图所在路径中不要有中文
字符),
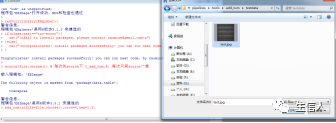
3、出现‘please click the first hole’时,点击第一个胶孔的位置(箭头指向,通常是Marker的位置),
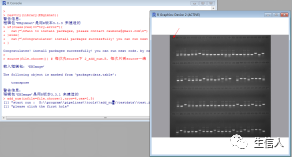
4、按照提示分别再继续点击第一排最后一个点样胶孔和第二、三、四排第一个胶孔(如下图),结束后会出现“add number successful”提示,表明运行成功,在胶图所在文件夹中会生成一个以“胶图名称_num”命名的标记好编号的胶图图片,大功告成!(注:这里是在Rstudio里运行该程序的方法,若在R程序中运行上述内容时,只需每次在胶图上点击胶孔后再点击一下R Console”命令行界面即可,其他内容与在Rstudio程序里运行方式完全相同。)
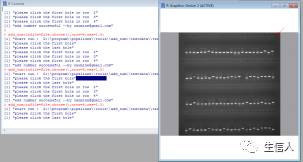
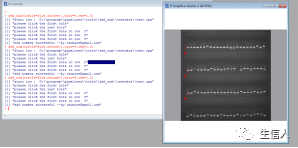
注意事项
1、在R和Rstudio里均可运行该程序,但在Rstudio比R里运行该程序更简单方便些。
2、用熟练后不看提示,出现图片后,依次点击下面的位置即可。
4. 总结
该程序结合人工识别能很方便快捷添加胶图编号,大大提高读带效率,并通过设置nrow, start, end 满足不同的需求,使用简单方便。
使用时请注意文件名不要有中文字符。
PS:该程序和教程由[email protected]原创,感谢其供稿。
七、电泳结果分析
PCR扩不出来由很多原因,每个细节都有可能导致扩不出来哦。首先重做一遍实验,并且设阳性对照,让同学帮忙看着,以防那个组分没加,检查PCR条件,并优化。如果还扩不出来,将PCR体系中的酶、dNTP、buffer全换新,并且换台PCR仪器试试。实在不行,重新设计引物,一般我们都在设计巨多条引物时(几百)才用primer5,你可以自己设计,按照这个网站http://www.idtdna.com/analyzer/Applications/OligoAnalyzer/计算Tm和hairpin的Tm(以小于30℃为佳),这个是国外非常常用的。
(一)产量低或无PCR产物。
1.模板不完整。通过电泳检测模板的质量。
2. 模板纯度低。对于DNA模板,其中含有的酚(phenol),乙二胺四乙二酸(EDTA), 蛋白质及高金属离子浓度(如Mg2+浓度)等污染物会影响酶的活性而影响PCR反应。可以通过再次沉淀然后70%乙醇洗涤来进一步纯化DNA。.
3.错误的引物设计。对于DNA模板的引物,可以使用相关软件进行设计。
4.酶用量不足或效率低。酶应贮存在-20℃冰箱,现用现拿,避免酶长期暴露在室温而失活,因此可以适当增加酶的用量,但酶用量过多会导致非特异性扩增。
5.引物用量不足或效率低。引物应贮存在-20℃,避免反复冻融。构建PCR体系时一定在冰上操作,所有试剂都应插在冰上。
6.Mg2+量不足。如果Mg2+浓度过低,则会导致PCR产量降低。由于Mg2+浓度能够受引物、dNTPs,模板的影响,故Mg2+浓度必须相应的进行调整以获得最大产量。若模板中含有EDTA等金属螯合剂时,必须相应提高Mg2+浓度(一分子的EDTA能够结合一分子的Mg2+)。在某些需要高浓度的dNTPs的PCR反应中,由于dNTPs能够结合 Mg2+形成复合体,故必须相应增加Mg2+浓度。
7.模板GC含量丰富。对于DNA模板,可以适当提高退火温度。对于RNA模板,若GC含量丰富或具有二级结构,则可以适当提高一下反转录温度。
(二)无特异性PCR产物。
1.错误的引物。重新设计引物。
2.在室温下建立PCR体系。当PCR体系在室温下建立时,Taq酶在建立的过程中表现为低的但是显著性的活性,结果PCR反应中会出现非特异性扩增。为了避免这种现象若使用Taq聚合酶,须在冰上进行操作。也可使用热启动酶,该酶在室温下没有活性,只有处在高温的PCR反应中才具有活性。
3. Mg2+浓度过高。如果Mg2+浓度过高,则会出现非特异性扩增。推荐的使用浓度为1—4mM。如果Taq Buffer 中含有KCl则起始的Mg2+浓度为1.5mM,如果Taq Buffer 中含有(NH4)2SO4,则Mg2+起始浓度为2mM。
4.模板量过高。在50ul体系中,对于质粒和片段DNA,最佳的模板用量为0.01—1ng,而对于基因组DNA则为0.1—1ug。过多的模板将导致非特异性扩增。
5.退火温度不合适。为提高特异性,也可在最初的5个循环使用Tm高的引物作为Tm,后面的循环使用Tm低的,即采用Touch down PCR。
(三)PCR产物序列错误。
1.低保真度的耐热DNA聚合酶。对于下游的一些反应例如克隆、定点突变需要高保真度的DNA聚合酶,如pfu Polymerase。
2.Mg2+浓度过高。Mg2+浓度过高,就会降低PCR的保真性。推荐的使用浓度为1—4mM。标准的dNTP浓度为0.】
2mM的PCR反应,如果Taq Buffer 中含有KCl则起始的Mg2+浓度为1.5mM,如果Taq Buffer 中含有(NH4)2SO4,则Mg2+起始浓度为2mM。
3.非最佳的模板量。在50ul体系中,对于质粒和片段DNA,最佳的模板用量为0.01—1ng,而对于基因组DNA则为0.1—1ug。如果模板量过低,则会降低PCR反应的精确性。
4.暴露于紫外线下。为避免紫外线的伤害而造成序列的突变,可采用以下两种方法一是在切胶的过程中用长波长的紫外线(360nm)二是需要回收的DNA在跑电泳时将同一样品分两道跑,一道跑大部分的DNA,另一道跑能够在紫外线下分辨的DNA的量即可,在电泳结束后将含量多的用刀切下,少的用于紫外线下确定位置,然后通过比较将含量多的相同位置的胶切下用于回收。
5.错误的引物设计.
DNA 模板:避免两对引物之间直接重复,碱基对互补或自我互补,防止产生不需要的产物。
RNA模板:为了避免对基因组DNA的扩增,设计引物时位置应设在内含子和外显子交界处。也可以使用DNase I从RNA中除去基因组DNA。
(四)PCR产物出现在阴性对照中。
1.提取RNA中含有基因组DNA的污染,故在反转录前用DNAase Ⅰ将DNA消化。
2.错误的引物设计。
DNA 模板:避免两对引物之间直接重复,碱基对互补或自我互补,防止产生不需要的产物。
RNA模板:为了避免对基因组DNA的扩增,设计引物时位置应设在内含子和外显子交界处。也可以使用DNase I从RNA中除去基因组DNA。
Refferences
1. Rand, K.N., Crystal Violet can be usedto Visualize DNABands during Gel Electrophoresis and to Improve Cloning Efficiency,Elsevier Trends Journals Technical Tips, Online,T40022, 1996.
2. Adkins, S., Burmeister, M., Visualization ofDNA in agarosegels and educational demonstrations, Anal Biochem., 240(1),17-23, 1996.3. Hu,G., DNA Cell Biol.,12, 763-770, 1993.
4. Longo, M.C., et al., Use of uracil DNAglycosylase to con-trol carry-over contamination in polymerase chainreac-tions, Gene 93, 125-8, 1990.
5.fermanent材料
八、在测序行业的主要应用
目前电泳技术在测序行业的应用主要有两个方面:一、纯化与检测DNA,在提取完DNA之后要进行跑胶,看提取DNA质量如何。二、样品测序之前,要对DNA进行打断成要建文库大小的片段,这里还需要筛选出相应长度的DNA片段。
九、新的电泳技术

新的电泳技术其实原理和上面的一样,不过是更准、更精确。比如:毛细电泳、脉冲电泳、双向蛋白电泳等。
光学图谱技术中采用的纳米空道电泳,通过电泳让DNA通过纳米孔道,在这个过程中实施拍照,获取其光学图谱。
第六章 Southern Blot
实验原理
Southern Blot用于凝胶电泳分离的酶切后DNA片段混合物中的特定性性片段(目标基因)的检测 。首先提取基因组DNA,然后酶切切成小片段,使用吸水纸将凝胶中的DNA通过虹吸作用被转移到尼龙膜上,然后用标记生物素的探针(核酸序列)与转移到膜上的DNA进行杂交,最后通过显色检测目标基因的存在性。Southern Blot主要用于检测目标基因的存在性及拷贝数多少,尤其用于检测转基因植物染色体基因组中是否已插入外源基因。
实验准备
1.固相支持介质
常用的有尼龙膜(nylon)和硝酸纤维素膜(NC)其特点如下:
(1)正负电荷尼龙膜是较理想的核酸固相支持物,有多种类型;硝酸纤维素膜是目前应用最广的一种固相支持物,价格最便宜。
(2)就结合能力而言:尼龙膜结合DNA和RNA能力可达480-600μg/cm2,可结合短至10bp的核酸片段;硝酸纤维素膜具有较强的吸附单链DNA和RNA能力,特别再高盐条件下其结合能力可达80-100μg/cm2,对于200bp的核酸片段结合能力不强;
注:硝酸纤维素膜在低盐环境下结合核酸能力不强,因此不适合用电转移法。
(3)就结合方式而言:硝酸纤维素膜靠疏水相互作用来吸附核酸,容易被洗脱。而尼龙膜则通过离子键结合,结合牢固,不容易洗脱。
(4)就温度适应性而言:尼龙膜经烘烤或紫外线照射后,核酸中的部分嘧啶碱基可与膜上的正电荷结合;硝酸纤维素膜依靠疏水性相互作用结合DNA,结合不牢固;
(5)就韧性而言:尼龙膜较强;硝酸纤维素膜较脆,易破碎;
(6)就重复性而言:尼龙膜可反复用于分子杂交,杂交后,探针分子可经碱变性被洗脱下来;硝酸纤维素膜不能重复使用。尽管如此,尼龙膜一般也不反复使用,避免可能的因素对结果产生影响
2..滤纸。购买大规格的滤纸。
3..托盘,玻璃板,直尺,铅笔,保鲜膜。
4.fermentas探针标记检测试剂盒,预杂交液二,及其它试剂(见试剂配制)
时间安排
第一天:基因组的提取及酶切:上午提取,下午检测,测浓度,晚上酶切。
第二天:上午检测酶切情况,并作浓缩,并跑上电泳;下午标记探针,配试剂,晚上转膜。
第三天:上午继续转膜,下午预杂交,杂交。
第四天:上午洗膜检测。
第五天:看结果。
实验步骤
本实验标准采用带正电荷的尼龙膜。
(一)DNA提取参见实验。注:提取完后,加入1/100体积的10mg/ml的RNAase,37℃处理40min,电泳检测后把所有离心管中的DNA合成一管然后用快速核酸检测仪测量DNA的浓度。
Southern杂交对基因组DNA的质量要求较高,用于Southern杂交的DNA必须长度完整,
大小在20kb-50kb之间,没有剪切或降解。
(二)DNA酶切:对于基因组的酶切一般选用多种限制性内切酶,既可以单酶切也可以双酶切或多酶切。选酶的原则是该酶在该基因内部无切点,且具有识别六个碱基序列的核酸内切酶(eg:EcoRⅠ,BamH Ⅰ,Hind Ⅲ)这样得到的酶切片段较大,易于吸附于膜上,不容易洗脱。同时应避免使用甲基化敏感的酶(eg:Pst Ⅰ,SalⅠ,XhoⅠ,ClaⅠ)酶切所需DNA的量一般为10μg~20μg,所需限制性内切酶量为10Unit酶/1μg
400μl的酶切反应体系:
10~30μg DNA
酶40ul(一般选取2—3种酶进行切割)
10×Buffer 40ul
BSA 40ul
无菌水至400ul
酶切体系大小的确定一般根据DNA量的多少确定,适当扩大酶切体系可以保证酶切完全。根据酶的使用说明书选择相应的酶切温度,酶切16h。取10ul电泳检测酶切效果。正常电泳图谱呈现一连续的弥散带,但有时还经常可见到一些明亮条带,这是重复序列而不是电泳假象。但是如果靠近点样孔附近有一条明亮的带,说明酶切不完全。如果泳道的边缘较亮,说明点样过大。如果泳道中间变宽,下段前缘变细小,说明是电压较高造成。如果消化不彻底,可延长反应时间,但是一般不能超过20h。如果延长时间不凑效,可采用补充酶液再消化的策略。结果图片如下:
注:
1. 建议使用TBE,缓冲能力强,产热量少。另外goldview要少加,如40ml胶加1ul即可。
2. 胶板用长的,以适合长时间电泳,保证各种不同长度大小的DNA片段能够更加有效的分离。
3.除酶切产物外,还应有marker,同时设置阴性(ck)、阳性对照(该基因的PCR产物或质粒),DNA。以对转膜杂交过程进行检测
4. 在进行大批量基因组DNA过夜酶切时,为避免酶长时间反应而导致活性的降低,最初加入的酶量应为所需量的1/2,酶切过夜后第二天早上再加入另一半,然后继续反应1小时即可实现完全酶切。
5.为了避免型号活性(识别位点错误),注意所加酶的用量不能超过酶切体系的10%即甘油含量不能超过5%。
6.甲基化酶能使DNA 不被相应的限制酶所切割,导致酶切失败。
(三)浓缩:加入1/10体积(40μl)3mol/L醋酸钠(pH 5.2)2倍体积的冷无水乙醇(或0.6~1倍体积的预冷异丙醇)-20℃沉淀DNA30min,12000r/min,离心10min。加入70%冷乙醇1ml 12000r/min,离心10min,洗涤沉淀,干燥,溶解于25μl 无菌水。
注:干燥时一定干燥至无酒精味为止,否则点样时sample may float out of well。
(四)低电压电泳:
1.关于酶切后电泳的点样。由于酶切产物经过浓缩,故浓度相当大,所以在点样时,上样缓冲液不应按照稀释至一倍即可,而应加大用量,否则将不能有效沉降DNA,而造成样品损失。另外要适当加厚一下胶,防止样品无法全部点入。
2.换用新的1×TAE或0.5×TBE电泳缓冲液和0.8%的琼脂糖凝胶以小于1V/cm(20V
左右)的电压电泳8—12h。电泳完成后,用凝胶成像系统拍照,若DNA没能有效的分离,可以将凝胶放入电泳槽继续电泳。
3.基因组样品上样时应使样品在点样孔中分布均匀后,再开始电泳,且最初电泳时应保持较高电压,以保证样品出孔。
(五)标记:修去凝胶边缘和加样孔上以及存在RNA的部分切下,并在Marker侧,切去一小三角,作为凝胶方位的标记。并用直尺测量胶的长宽,确定其大小。
6,7,8步操作时一定要小心,保持胶的完整性。
6.脱嘌呤处理:对于大于20kb的条带,可采用将凝胶置于0.2mol/L的HCl(无菌水配制)中摇晃之溴酚蓝变黄(一般5—10min),二甲苯青变成黄色/绿色后,迅速将HCl倒入废液缸,无菌去离子水洗涤数次。(强酸导致DNA脱嘌呤,能够减小DNA的大小而有利于转膜。但是,过度会导致DNA断裂成较小片段。)注:凝胶是否需要脱嘌呤处理取决于DNA片段的大小,一般可根据marker判断,如果序列>20kb,则需要脱嘌呤处理,从而保证转膜的效率,如果序列<20kb则不必进行脱嘌呤处理,否则可能出现过嘌呤,导致DNA片段太小而不能有效结合到膜上。
注:如果实验转膜过程中同时含有大于 15kb的DNA(通常为基因组)和小片段DNA(通常是基因组酶切产物),那么在转移的过程中倾斜容器,仅使凝胶上半部分浸泡于盐酸,这样既可以提高大片断 DNA 的转移效率,又不会打碎小片段DNA。
7.变性处理:加入数倍体积的变性液,每次15min,摇床处理两次,然后无菌水冲洗。
8.中和处理:加入数倍体积的中和液,每次15min,摇床处理两次,然后无菌水冲洗。
从DNA固定于膜上到其后的杂交,这一系列步骤的顺序取决于膜的种类、转移的方法以及固定的方法。在碱性缓冲液中,DNA共价结合在带正电荷的尼龙膜上,因此杂交前不需要固定。在中性缓冲液中转移至不带电荷或带正电荷的尼龙膜上需要加热固定或紫外交联。本文以中性缓冲液为例,讲述。
9.转膜准备:
①、切一张每边比凝胶大1mm的尼龙膜,并用干净的解剖刀切下膜的一角,与凝胶所切下的角一致。同时剪10张同样大小的滤纸。(注意:需带一次性PE手套和钝头镊子,沾有油污的膜不易浸湿。)
②将尼龙膜飘浮于盛有无菌去离子水的平皿中,直到其完全浸湿后(一般5min),将其浸入适20×SSC中30min,。(注意:膜的浸湿完全与否,对DNA的转移至关重要。)而将10张滤纸浸泡在2×SSC中。
③将8张滤纸放在一片玻璃板上,形成比凝胶长且宽的支持物。将支持物放入一个大的托盘中,吸水纸两端从板边缘垂下。
④托盘中加入适当的20×SSC,直到液面几乎与支持物表面平齐。当支持物上的滤纸完全浸湿后,用玻璃棒赶走吸水纸下的气泡。
⑤将凝胶从中和液中取出,倒转使原来的底面向上。放在支持物上(使凝胶尽量与底层吸水滤纸的边同样长或略短,见图)。用玻璃棒赶走凝胶与吸水纸间的气泡。
⑥用保鲜膜包绕凝胶四周,不要覆盖有DNA的凝胶部分,以屏蔽转移缓冲液从凝胶周围短路流入吸水纸。(注:根据测量的凝胶的长度画出凝胶形状,然后在此基础上再画出一个每一边都比前面略小的图形。最后将保鲜膜放在这张纸的上面,然后象图2那样将保鲜膜固定,用刀片沿最后画的图形的痕迹将保鲜膜切下。然后这一保鲜膜既可用于封闭凝胶。)
⑦用适当的20×SSC溶液将凝胶湿润。将湿润的尼龙膜放置于凝胶上并使两者的切角相重叠。为避免产生气泡,当先使膜的一角与凝胶接触再缓慢的将膜放到凝胶上。膜的一边应恰好超过凝胶上部加样孔一线的边缘。膜一旦放在凝胶上就不要再移动了。
⑧在尼龙膜上放置10张与膜大小相同的滤纸,叠多叠卫生纸,将其放在滤纸之上,在卫生纸顶部放一400g重物压实。注:不可压过重的重物,因为这样可能会使胶伸展弯曲变扁,从而使最终长度的判断出现困难,且条带变粗。
⑨转移8~24h,并每隔2—3h更换浸湿的纸巾。尽量避免纸巾都被缓冲液浸湿。(对于大于16kb的24h以上)
⑩除去凝胶上的卫生纸和滤纸,翻转凝胶以及与之接触的尼龙膜,凝胶向上平放于干燥的吸水纸上。用极软的铅笔在尼龙膜上标记记加样孔的位置。
11将凝胶从膜上剥离,弃去凝胶。
13、紫外反射透射仪检测凝胶上DNA的残留量,用以确定转移效率。
10.DNA的固定:
1、将膜浸入6×SSC,室温放置5min,以除去粘附在膜上的凝胶碎片。
2、①真空烘烤固定:将膜从6×SSC中取出,并使多余的液体流净。将膜放在滤纸上室温晾干30min,将膜夹在两张干燥的滤纸中间,真空炉中85℃烘烤3h。(也可直接在杂交炉中烘烤)
②紫外照射交联:将潮湿的膜放在一张干燥的吸水纸上。254nm的紫外线照射使DNA交联到膜上。(其原理是紫外线照射使DNA中的一小部分胸腺嘧啶残基与膜表面带正电荷的氨基基团之间形成共价交联,但过量照射将导致大部分的胸腺嘧啶共价结合而降低杂交信号。同时,紫外线照射可能增强某些带正电荷的尼龙膜的杂交信号背景,因而应当使膜上带有DNA的一面朝向紫外。建议对潮湿的膜采用总量1.5J/cm2,干燥的膜采用0.15J/ cm2。)
3、对于不立即用于杂交的膜都应充分干燥,松弛的包于铝箔纸或吸水纸中,室温下最好保存在真空条件。或待膜冷却后,用保鲜膜包好以后保存在4℃备用。
限于篇幅更多介绍参见:
Southern Blot杂交
第七章 基本实验操作
分子生物学基本实验操作
欢迎关注生信人











