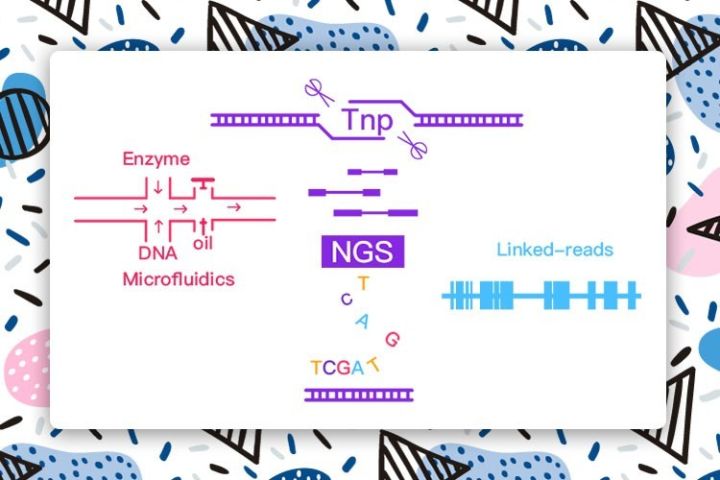
测序是基因编辑的前提,测序在诊而编辑在于治。与CRISPR/Cas9作为防御武器不一样,转座酶/Tn5更像是物种交战后在基因组上留下的残骸;Cas9和Tn5都具有核酸酶活性,不过前者追求特异性,而后者追求随机性。二者都具备简捷、高效和灵活性强的特点,这也使得它们在各自的领域发挥着举足轻重的作用。转座酶(Tnp)就像是基因测序的魔剪。
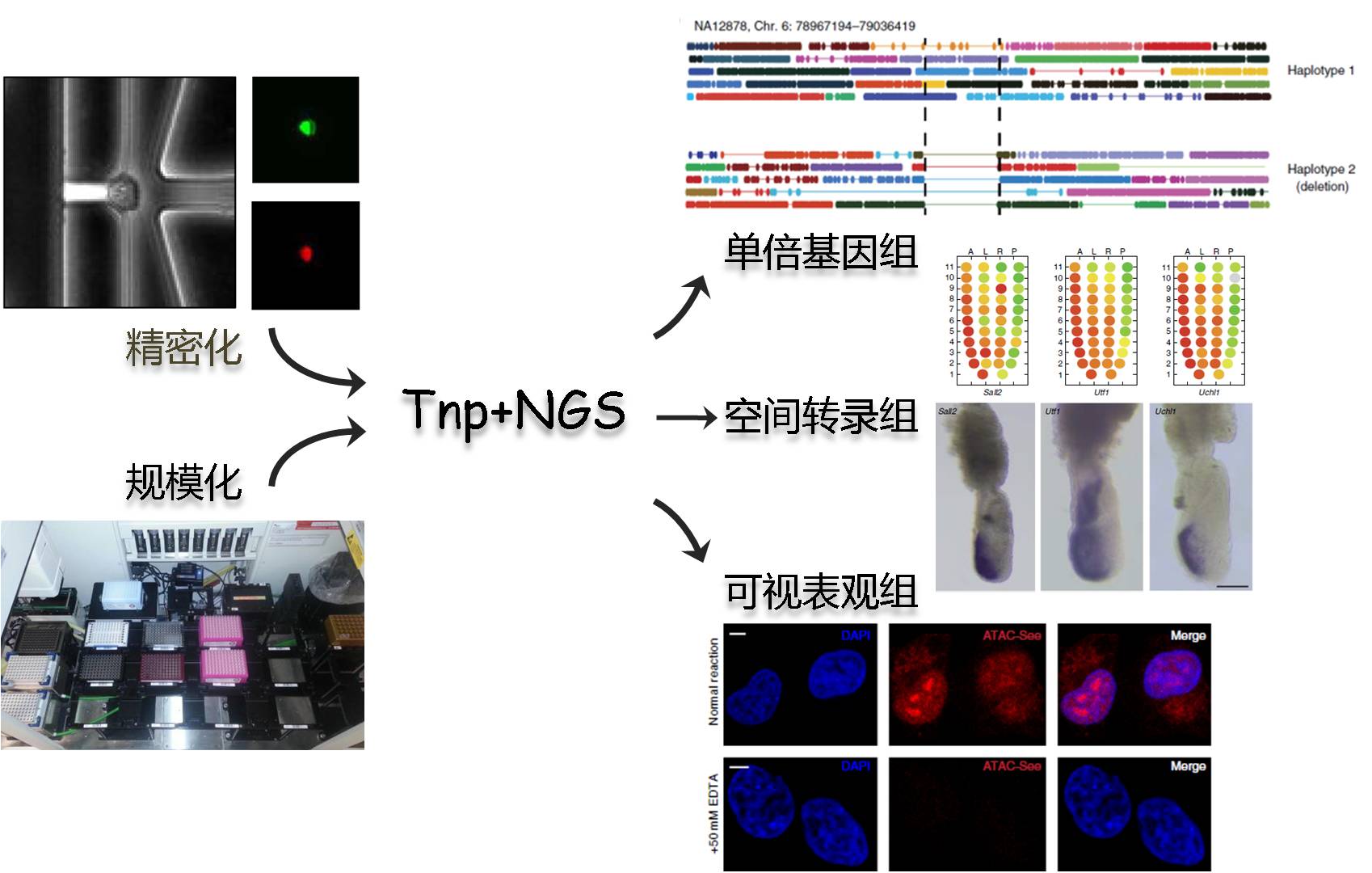
图1:转座酶助力二代测序应用[1-5]
在NGS高速发展的10年间,测序及相关技术逐步成熟,并朝着精密化和规模化发展。科学家不再满足于平均值,他们更希望从单细胞、单分子层面解读遗传信息;面临肿瘤的异质性,规模化测序显得尤为重要。面对这些挑战,转座酶帮助NGS实现了多个技术突破,加速了相关领域的研究。
研究人员发现,整个转座子序列并不是转座必须的,只需转座子的末端核心序列,转座酶便能将该部分序列插入并连接至基因组内;根据这个原理,将测序接头序列加入末端核心序列中,可简捷地引入测序接头,完成文库构建。
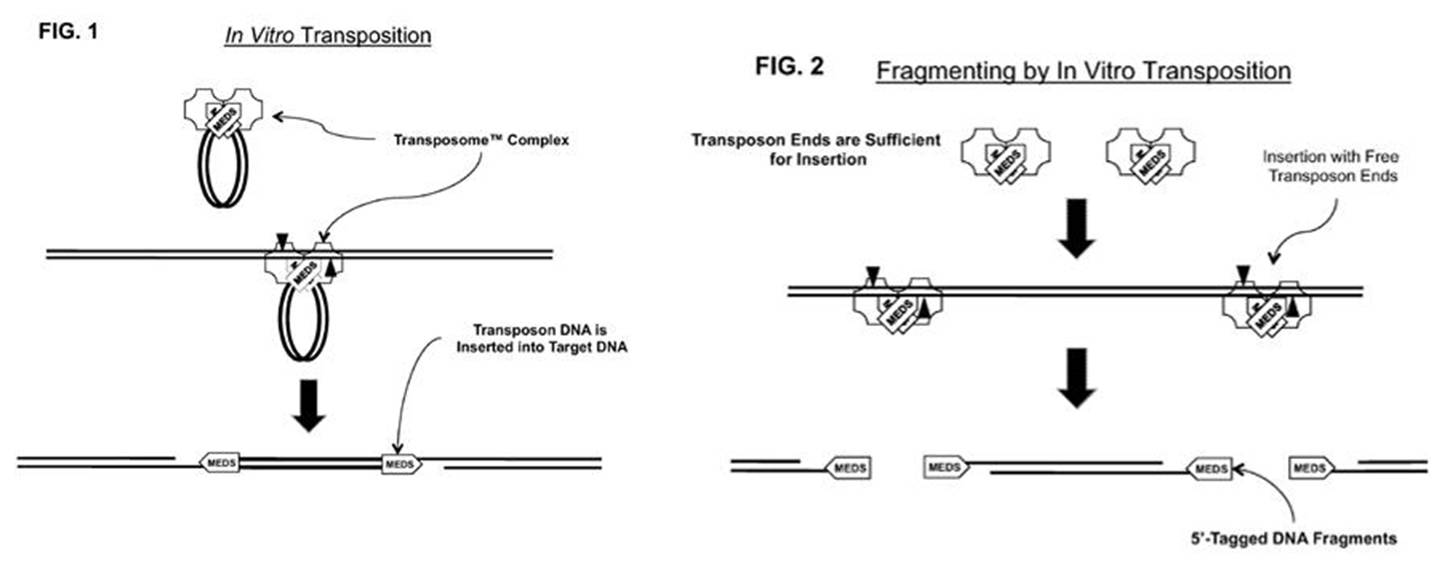
图2:转座酶改造以及建库原理[6]
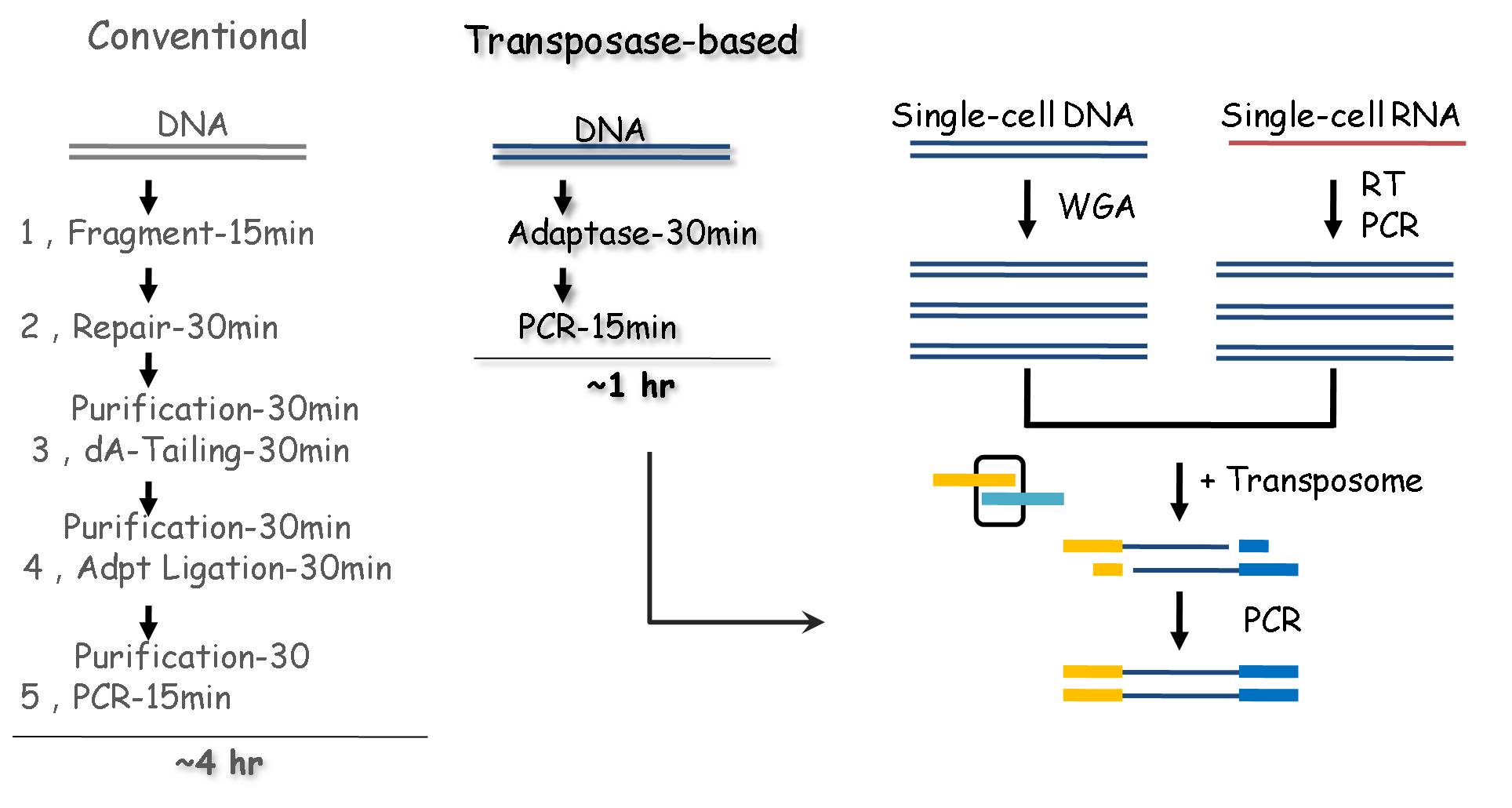
图3:转座酶建库流程以及应用于单细胞测序
1. 简捷
转座酶同时具有切割和连接两种活性,且其工作缓冲液与PCR兼容,可将繁复的NGS文库构建“5合1”,极大简化了测序前处理。
2. 高效
转座酶也是低起始量、单细胞DNA/RNA建库的优选方案;同时具备简便、高效两个特点是转座酶实现自动化、规模化测序的先决条件。
3. 灵活
转座酶的接头序列可操作性强,研究者可根据实验目的进行改造,是一种平台型建库酶。例如,加入生物素修饰研究基因组结构变异、加入荧光基团对表观基因组调控进行可视化研究等等。
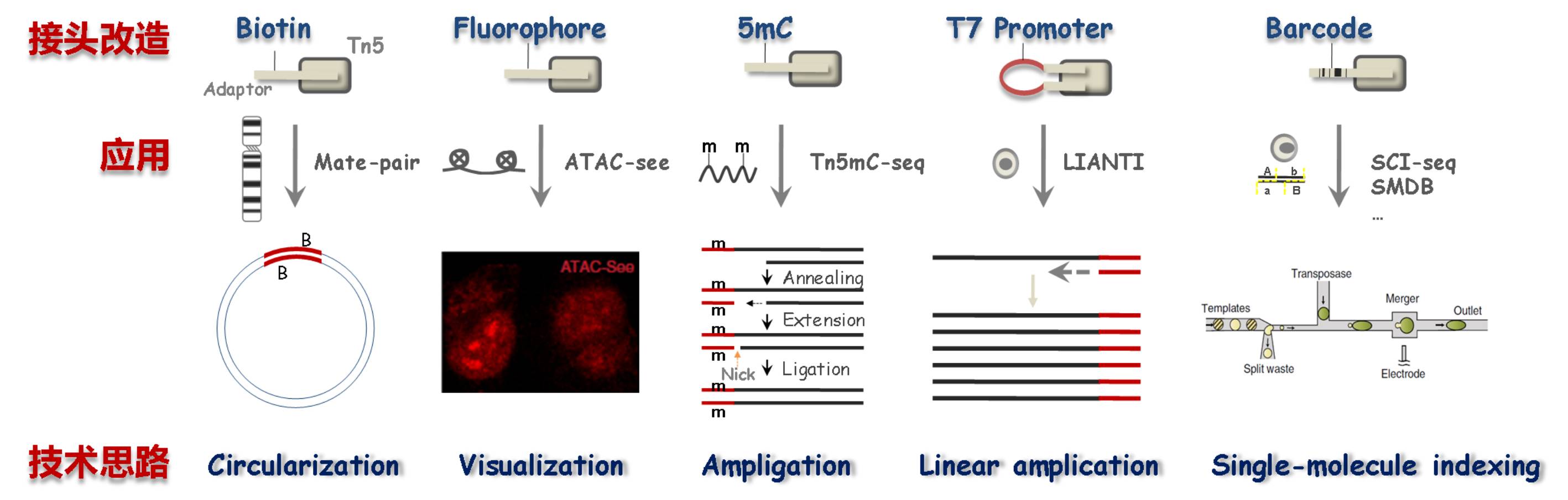
图4:转座酶接头改造[5][7]
1. 扬长避短-结构变异检测
在短读长的限制下,二代测序并不能直接对长片段进行测序,Mate-pair测序技术使用转座酶接头的核心序列标记DNA片段的两端,成环后只对标记区序列测序便能探测基因组序列的大致位置关系,在医学研究中多用于寻找结构变异的断裂点。
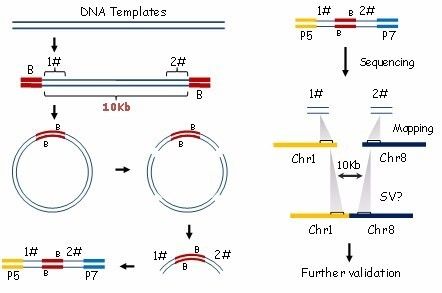
图5:Mate-pair测序技术流程图
2. 扬长避短-单倍体分型
同样受限于短读长,单倍体分型一直是二代测序应用中的一个薄弱环节,利用Barcode标记的转座酶接头结合微滴微流控技术,可较为容易的使测序分辨率提升至单倍体水平。
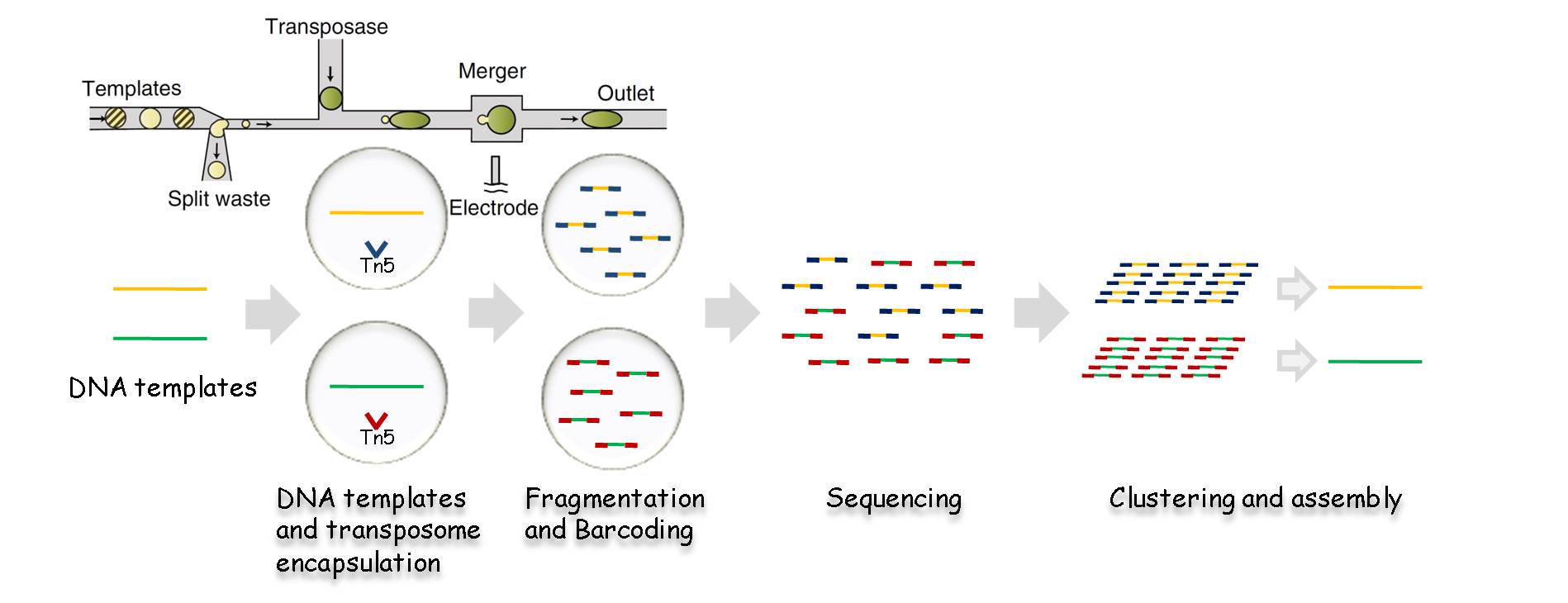
图6:基于微流控和Barcoding技术的单倍体分辨率测序流程[7]
3. 简化表观基因组研究
ATAC-seq利用转座酶代替DNase I核酸酶分析染色质可接近性,更加简便、投入量更低、通量更高;在转座酶接头上加入荧光基团后,ATAC-see将影像学和测序技术结合起来,能够对表观基因组的时空结构进行更为立体的解析[5]。
4. 提高单细胞DNA测序分辨率
LIANTI技术利用对称型接头提高转座酶模板利用率,并使用T7 promoter进行线性扩增降低偏好和PCR错误,可大幅提升单细胞DNA测序的CNV分辨率和碱基准确性。
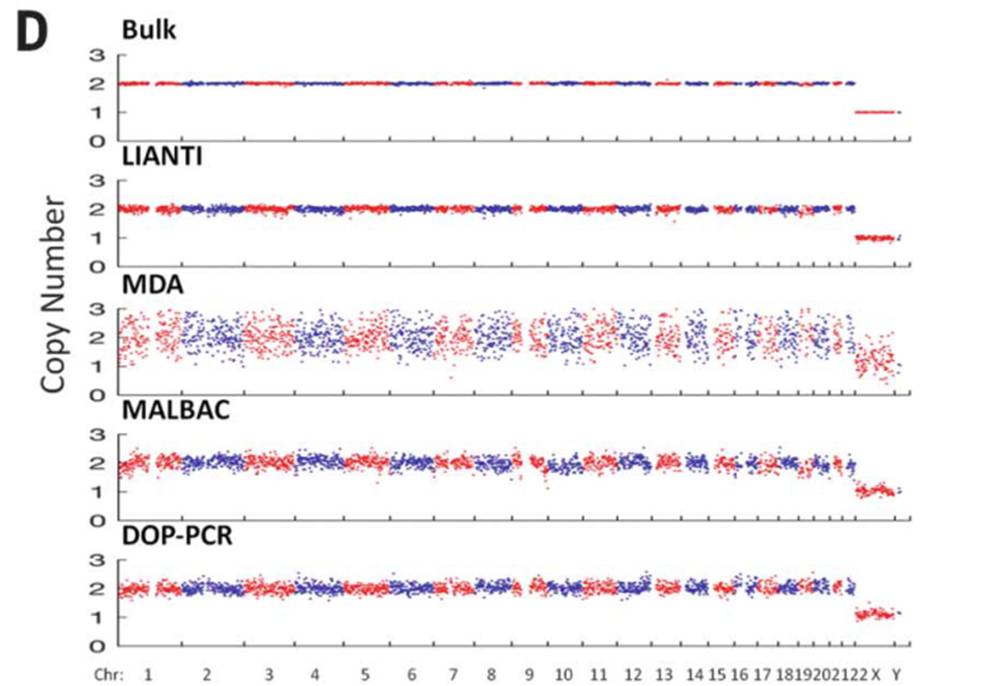
图6:LIANTI与其他单细胞技术的CNV分析比较[8]
5. 实现单细胞规模化测序
借助转座酶简便、高效的优势,研究人员已经开发出多种基于转座酶的单细胞DNA/RNA的规模化测序技术。比如,SCI-seq可同时检测上千个单细胞的拷贝数变异(5395-可分析数量/16698-建库数量),对于肿瘤异质性研究具有重要价值[9]。
1. DNA纯度
来自ATAC-seq技术的温馨提示,DNA结合蛋白会抑制转座酶对DNA的均匀插入,降低覆盖均一性;如果发现如下图所示的覆盖度分布,那么这个WGS很可能已被转化为ATAC-seq。
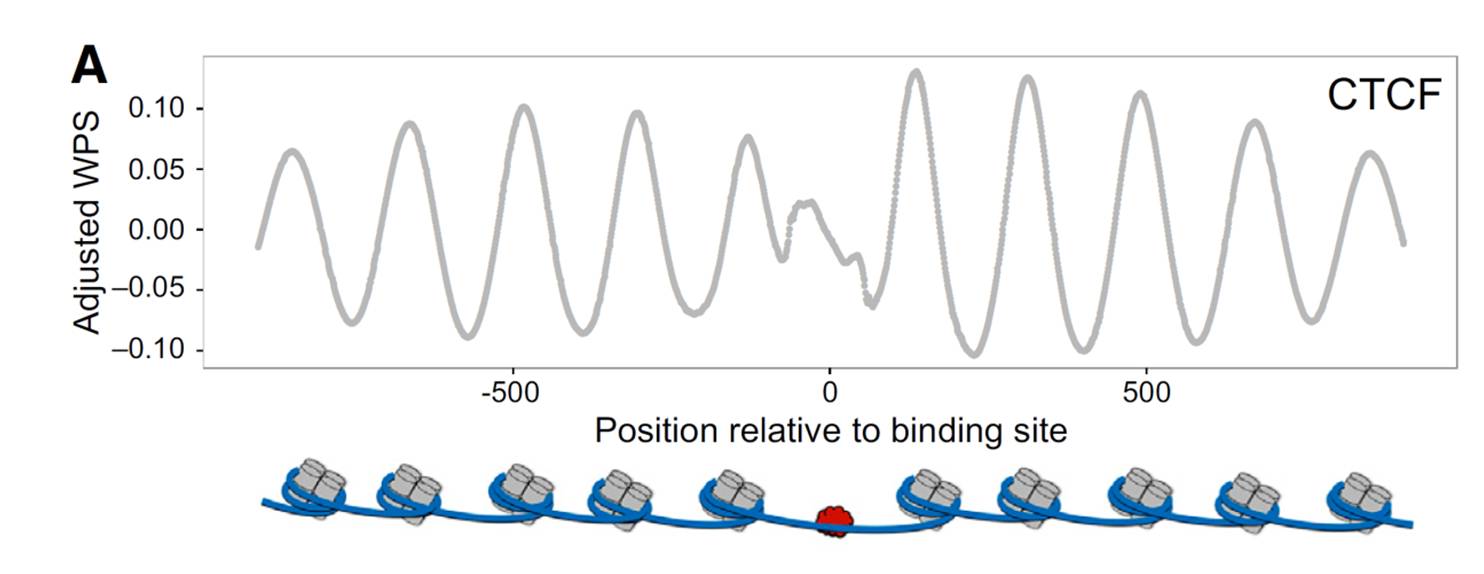
图:测序覆盖度与DNA结合蛋白的分布(10)
2. 偏好性
虽然Tn5转座酶已经被改造优化,但其插入位点序列的偏好性依然存在,请不要随意使用转座酶对某个物种进行测序。
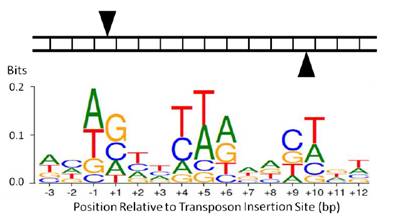
图:Tn5插入位点序列偏好[8]
3. 文库大小
转座酶文库大小对DNA投入量敏感,实验室在开展转座酶建库前,需要仔细摸索DNA投入量和转座酶的比例。需要注意的是,该特性也让大规模单分子转座酶建库更具挑战。
4. 对称型接头
利用对称性接头可以提高转座酶的模板利用率,上文提到的单细胞测序技术LIANTI使用就是对称性接头,其作用与线性扩增同样重要。
5. Tn5与DNA结合紧密
选择何种方式分离Tn5和DNA对后续建库效率影响较大,此外,该特性可用于不依赖微流控设备的单分子测序[11]。
容易想到的是,转座酶建库能够满足有短周期需求样本的检测,例如应用于CNV的快速检测;根据LIANTI的表现,对于提高PGS精确性值得期待;在液态活检或是NIPT方面,由于cfDNA都是~170bp的短片段,并不特别适合于转座酶建库。
发展之初,起始量过高,建库流程繁琐限制了NGS的应用,转座酶建库技术很好的解决了这些问题;随后,其他低起始量建库技术逐渐成熟,其流程也相对简单,转座酶的主要优势被削弱;随着NGS应用的深入,人们对数据质量的要求越来越高,不仅仅是完成测序,而且要测得更好,所以转座酶的偏好问题就被凸显出来,权衡利弊后,转座酶不再是第一选择。
在NGS平台上利用转座酶对长片段测序是短读长的权宜之计,相信三代技术成熟后能更好的解决这个问题。
虽然优势被削弱也有偏好性的弊端,但转座酶建库仍然是最为简单的NGS建库流程之一,更具流线型的两步建库是大规模测序时的首选方案,所以,魔剪转座酶仍然会在二代测序领域发挥重要作用。
参考文献:
1. Single-cell chromatin accessibility reveals principles of regulatory variation.
2. Separation and parallel sequencing of the genomes and transcriptomes of single cells using G&T-seq.
3. Haplotyping germline and cancer genomes with high-throughput linked-read sequencing.
4. Spatial transcriptomic analysis of cryosectioned tissue samples with Geo-seq.
5. ATAC-see reveals the accessible genome by transposase-mediated imaging and sequencing.
6. Transposon end compositions and methods for modifying nucleic acids.
7. Droplet barcoding for massively parallel single-molecule deep sequencing.
8. Single-cell whole-genome analyses by Linear Amplification via Transposon Insertion (LIANTI).
9. Sequencing thousands of single-cell genomes with combinatorial indexing.
10. Cell-free DNA comprises an in vivo nucleosome footprint that informs its tssues-of-origin.
11. Haplotype phasing of whole human genomes using bead-based barcode partitioning in a single tube.
· END ·











