生物制药行业在最近20年发展迅速,特别是基于哺乳动物细胞表达的产品市场份额不断扩大。哺乳动物细胞(如CHO)的生产工艺开发过程通常包括分子构建、稳定细胞株筛选、上游发酵工艺开发、下游纯化工艺开发以及制剂工艺的开发。在这些阶段中,分子构建和稳定细胞株筛选做为一个产品研发的起始部分,显得极为重要。一株稳定高产的工程细胞株不仅能显著增加单位体积产量,降低产品的生产成本,并且还可以降低下游纯化过程的复杂度,确保获得安全,高品质的生物制品。
动物细胞是生产治疗性重组蛋白的强有力工具。为与小分子药物区别开来,治疗性蛋白、疫苗以及细胞治疗产品统称为生物制品。宿主细胞的选择和改造对于提高产品的产量和质量来说至关重要。因此必须对细胞进行筛选,选出具有高表达量和良好生长特性的细胞。在此我们主要讨论适用于大规模生产重组蛋白的细胞株的筛选。
工业上重组蛋白的制备主要有两种方式:瞬时转染和稳定转染。瞬时转染经常用于制备少量蛋白(多达克级)用于药物开发早期的蛋白活性或动物实验。瞬时转染并不产生克隆细胞株,它只是暂时将编码外源蛋白的质粒转导入已有的细胞株里面(比如HEK293或COS细胞)。随着细胞的分裂外源基因不断丢失,蛋白产量较低,一般在1-100mg/L。近年来有报道称HEK293细胞中瞬时表达产量可达1g/L。
与瞬时转染不同,稳定细胞株是为工业化生产治疗性蛋白而构建的。稳定细胞株需要具备在不同的时间,不同的场地以及不同的批次间生产相同质量产品的能力。在选定生产用细胞株之后,需要建立主细胞库(mater cell bank)。从主细胞库复苏后,扩增建立工作细胞库(working cell bank),工作细胞库用于生产。细胞库通常保存在液氮中,需要维持整个产品的生命周期。在构建生产用细胞株的过程中,宿主细胞和表达载体至关重要。
CHO和骨髓瘤细胞是构建稳定细胞株最常用的两种宿主细胞。基本上用这两类细胞生产的治疗性蛋白都能分泌到细胞外,从而可以从细胞培养液中收获产物。CHO细胞属于成纤维细胞,是一种非分泌型细胞,它本身很少分泌内源蛋白。
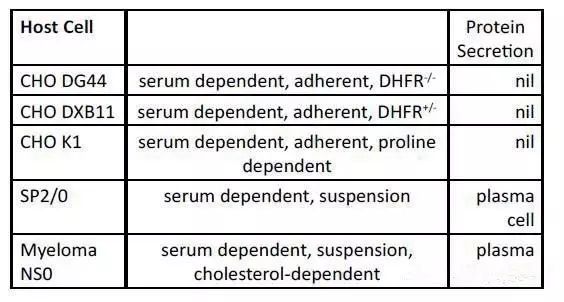
科研人员在蛋白组学和转录组学水平上研究了B细胞变成浆细胞所发生的生理变化。细胞转变提高了能量代谢水平,改善了蛋白分泌和糖基化能力,同时增加了氧化还原水平用来应对活性氧的影响。B细胞或浆细胞的基因组中只有一个功能性的免疫球蛋白基因。在B细胞成熟的过程中,二倍体细胞中其中一个等位基因发生失活。但是,仅仅是免疫球蛋白基因的一个拷贝足以使浆细胞成为一个高产的分泌细胞。因此,只需将目的基因的一个拷贝整合到杂交瘤细胞基因组适合的位置就可以使其成为分泌外源蛋白的强大细胞。与瘤细胞不同,CHO必须经过改造后才能提升蛋白表达能力。
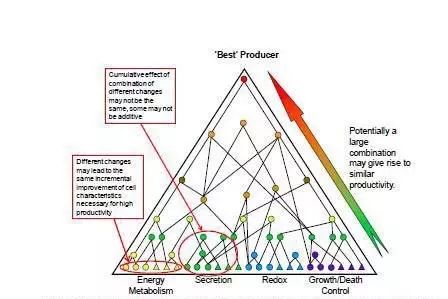
除了蛋白表达能力之外,优秀的细胞株还应具备良好的生长和代谢特性。在过去的十多年里,大量转录组学和蛋白组学的研究致力于揭示高产细胞株的特性。人们逐渐意识到没有那一种主要的调控因素能使CHO或NS0来源的重组细胞变成高产细胞株。高产细胞株的形成涉及细胞内许多通路的改变,比如代谢、分泌、氧化还原平衡和生长/死亡调控;更可能的是大量基因表达水平的微小变化,而不是几个主要节点的大变化。
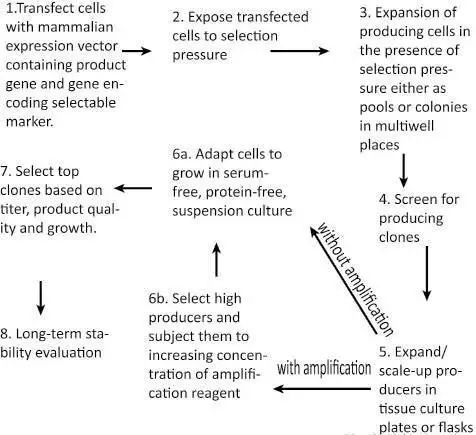
转染-筛选-扩增-单克隆化-筛选-驯化
首先,通过质粒将目的蛋白的编码基因导入到细胞内。质粒上除了目的基因外还带有抗性基因,比如抗生素抗性基因。这样在转染后就可以利用选择压力富集整合质粒的细胞。在哺乳动物细胞中质粒不能进行复制,未整合到基因组中的质粒随着细胞分裂逐渐丢失。在一段时间的选择压力后,所有存活下来的细胞基因组中都整合了外源质粒。
获得了一系列整合了外源基因的稳定细胞后,接下来就需要从中筛选出表达量最高的克隆。最常用的方法是检测96孔细胞培养板中蛋白的浓度;另外一个方法是让细胞在软琼脂上生长形成集落,然后利用原位免疫沉淀技术获得高产细胞。近年来,发展起来的高通量自动化筛选技术也被受关注。
通常使用的转染方法主要有DNA-磷酸钙共沉淀法,电击法,脂质体法。
质粒DNA的基本元件
外源基因是以质粒DNA的形式进入动物细胞的,同时质粒DNA上带有提高外源基因转录和翻译水平的元件。尽管病毒来源的载体和细菌来源的载体都被用来将外源基因导入动物细胞,但是工业上在构建细胞株的过程中极少用到病毒来源的载体。
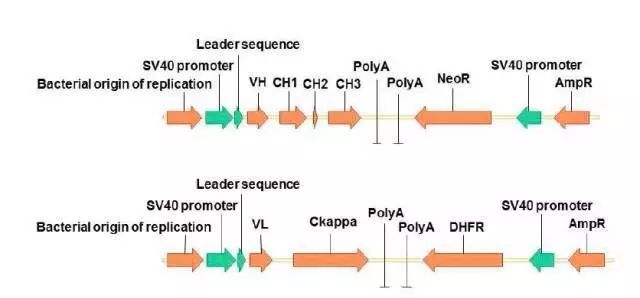
目的基因可以由组成型,诱导型或条件型启动子进行启动转录。在细胞分化和发育的研究中经常用到条件型启动子,它可以由某些特定因素引发,导致蛋白表达,影响细胞的分化。但是在重组蛋白的生产中,绝大多数表达载体使用强组成型的的启动子。通常病毒来源的启动子,如SV40晚期启动子和CMV启动子,应用最为广泛。近年来,CHO来源的EF-1和GAPDH的启动子也被应用到蛋白生产中。
为了提高转录水平,除了使用强的启动子外,还可以在目的基因中插入内含子。通常可以看到在目的蛋白的基因中含有至少一个内含子。此外,密码子优化可以提高基因的翻译水平。表达载体除了包含启动子、增强子和目的基因外,还需要有筛选标记基因,甚至,扩增标记基因。
尽管转染后有大量的质粒进入细胞,但是只有极少数能够进入细胞核,进一步整合到基因组中。细胞中只有整合的质粒才能复制,未整合的质粒逐渐被降级或丢失。转染后至少有一个筛选标记基因(抗性基因)整合到基因组并成功表达的细胞在抗性的压力下才能存活。
筛选标记可以分为显性的和隐形的。隐形筛选标记是细胞内固有的基因,缺失时影响细胞生长(如DHFR)。通过引入外源补偿基因可以克服这类缺陷。比如,CHO DG44 细胞是DHFR基因双敲除的细胞株,需要在培养基中添加HT才能正常生长。通过转染使其获得功能性的DHFR基因,就能使它在不含HT的培养基中也能生长。相反,显性筛选标记(一般为抗生素)对细胞具有杀伤作用。通过向细胞中引入抗性基因可以赋予细胞对抗显性筛选标记的能力。每一个抗性基因编码一种酶,可以破坏显性筛选标记的化学结构。下表(table2)中显示的是常用的显性筛选标记和抗性基因等信息。
参与使抗生素失活的酶(由抗性基因表达)只有在细胞内才有活性,失活过程中的磷酸化和乙酰化反应需要ATP和乙酰基团等底物,因此抗生素的失活必须在细胞内部才能进行。另一方面,蛋白酶即使在细胞死后释放到培养基中仍有活性。因此,在细胞株筛选过程中,筛选试剂(如抗生素)的浓度随时间逐渐降低,降低的速率取决于转染后细胞的密度。所以,最优的筛选试剂浓度不止取决于细胞系,也取决于细胞的密度。单克隆筛选和细胞群体“Pool”筛选在抗性浓度的使用上可能大不相同。
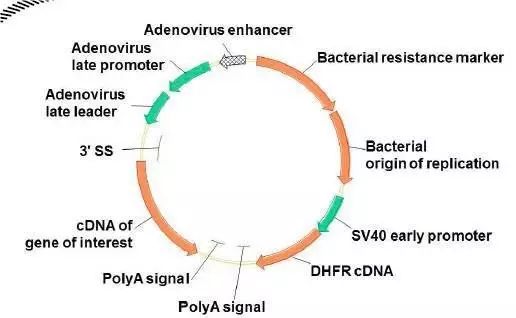
哺乳动物细胞中最常用的两种基因扩增系统是:二氢叶酸还原酶(DHFR)和谷氨酰胺合成酶(GS)系统。DHFR是催化二氢叶酸还原成四氢叶酸的酶,四氢叶酸是甘氨酸、胸苷一磷酸和嘌呤生物合成所必需的。氨甲喋呤(MTX)是叶酸的类似物,可以与DHFR结合并抑制其活性。从而使细胞在缺乏胸苷和嘌呤的培养基中死亡。
当细胞在MTX的压力下生长时,只有DHFR基因扩增并高效表达的群体才能存活下来。DHFR基因扩增可以带动插入位点附近10-10,000kb的DNA序列一起扩增。因此与DHFR邻近的外源基因也随之扩增。
扩增可以使一步完成也可以是分几步完成。随着MTX浓度的升高,存活下来的细胞DHFR扩增程度越高。能耐受高浓度MTX压力的细胞,可能含有几千个拷贝的DHFR基因。
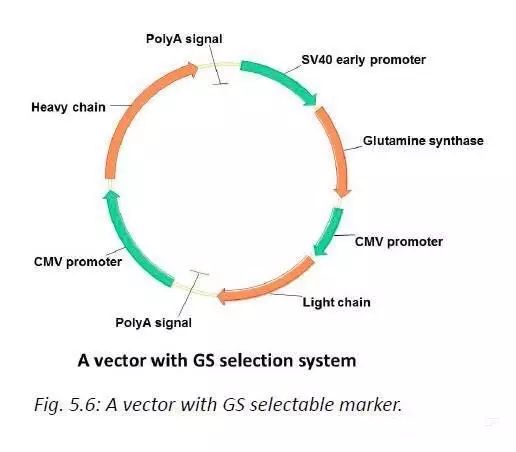
GS筛选系统基于细胞中的谷氨酰胺合成酶可以利用谷氨酸和氨合成谷氨酰胺。绝大多数哺乳动物细胞内源的谷氨酰胺合成酶活性很低,需要在培养基中额外添加谷氨酰胺细胞才能生长。GS筛选系统的载体含有谷氨酰胺合成酶和外源基因,因此可以在不含谷氨酰胺的培养基中进行筛选。通常用比较弱的启动子,如SV40启动子来启动GS基因。在高浓度的谷氨酰胺合成酶抑制剂(MSX)的作用下,可以筛选得到基因高度扩增的细胞。
为实现外源基因的高效表达,基于上述筛选系统人们开发出了更为复杂有效的筛选途径。比如把DHFR与G418抗性基因相连,转染后加G418压力,这时只有质粒载体整合到转录活跃区的细胞才能表达足够高的潮霉素抗性,抵抗G418的压力从而存活下来。这时筛选出的克隆只含有外源基因的少数几个拷贝。接着通过MTX的扩增作用,使整合基因在细胞基因组中进一步扩增,从而实现外源基因的高表达。
通过为期一到两周的基因扩增过程,筛选试剂浓度相应降低。在较低的筛选压力下,外源基因的拷贝数可能减少,导致转录水平和蛋白产量的下降。拷贝数的丢失与外源基因整合到染色体上的位置有关,位于染色体两臂末端的基因更倾向于丢失。通常在降低筛选压力后,细胞株先是经历一个外源基因拷贝数和表达量迅速下降的过程,随后便趋向于稳定。所以,通常需要维持一个合适的选择压力来防止细胞不利突变的发生,影响拷贝数或引起细胞生长速率的变化。
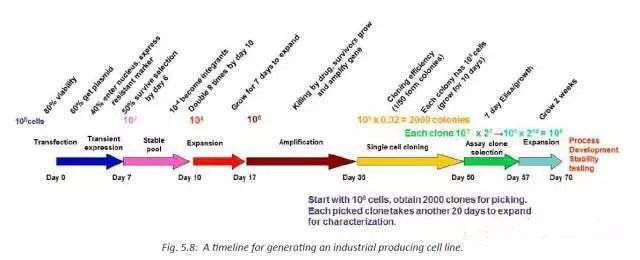
确定生产用细胞株后,需要尽快使其适用生产规模下的培养条件。
1.从细胞复苏(2×108个细胞)到生产(20m3)细胞至少需要分裂16次;
2.稳定性试验至少测试细胞分裂40次;