抗体广泛存在于生物医学研究、诊断和治疗的发展中,且需求量大。然而,满足全球市场日益增长的抗体需求量是一个巨大的挑战。为了维持抗体的供求平衡,科学家们发明了更好的方法来优化抗体的表达,以达到增加抗体的单体积表达量并减少生产成本。由于新的克隆方法、基因改造技术以及一系列的细胞和载体工程技术,加上发酵技术上的改进,这些都使抗体表达大大得益。
抗体表达取决于多种因素的复杂相互作用,且可能需要这些因素在不同水平进行微调以获得最大产量。然而,每个抗体都是独特的,在优化相关的表达要素时需要个体化的考虑。这篇综述旨在提供关于一些能够提高抗体表达的最前沿的方法的全面概述,例如宿主细胞选择、工程菌改造、密码子优化、基因优化、载体修饰和工艺优化等。
1. 前言
2. 宿主细胞的选择
3. 工程菌改造
4. 密码子、基因的优化
5. 抗体形式
6. 载体修饰
7. 优化表达参数
8. 总结
生物研究上的突破性技术--杂交瘤技术--的出现,让科学家们能够生产特异性的抗体,并且基因工程技术使生产全新的抗体成为可能,这些被修饰的抗体其优越性甚至超过自然的抗体,使得抗体有全新的应用。重组抗体克服了产量上的瓶颈并成为生物制药行业发展最快的部门。
强大的基因工程技术的发展促进了从单克隆抗体到重组抗体的初步转变。这些技术提供了操纵抗体基因的能力,使其可以产生所需的抗体框架、结合和效应子特性,并允许轻易加入标签和融合蛋白来协助检测、结合、纯化和增加血清半衰期。
高通量筛选平台的可用性、易于遗传物质的易于保存、抗体稳定的回收率和重复性、在廉价微生物宿主中的易于生产以及专利方面均增加了重组抗体的商业价值。
抗体表达是始于从DNA到mRNA的转录并在mRNA翻译成蛋白时达到高潮的多步骤过程(如图1所示)。抗体通常都具有复杂的结构,需要复杂的细胞器来完成正确的表达、折叠和加工。抗体的结构变异性为了满足不断增长的生物医学需求已经产生了复杂的抗体结构(如图2所示):分子量范围从12 kDa到150 kDa的一、二、三和四价结合能力。抗体的大小、序列、形式、二硫键数量和翻译后修饰都会影响其表达。这些优异抗体设计的最优功能表达和纯化是决定它们现实应用命运的关键因素。比如:双特异性抗体曾因低表达量和产品不稳定经历了很长一段时间的休眠。直到最近,双特异性抗体因着现代分子生物学工具的帮助,得以解决低表达量的问题,从而再次复活。大部分重组抗体是基于免疫球蛋白结构的抗原结合部位,而这些部位对构象改变敏感,并可能导致表达抗体的聚集,从而使其不适用于应用。没有一个通用平台可保证收获的重组抗体的良好功能,故而谨慎优化每个抗体的表达和纯化条件(如图3所示)以达到最大收益就显得非常重要。本综述旨在总结探索优化抗体表达的不同策略和最新趋势。
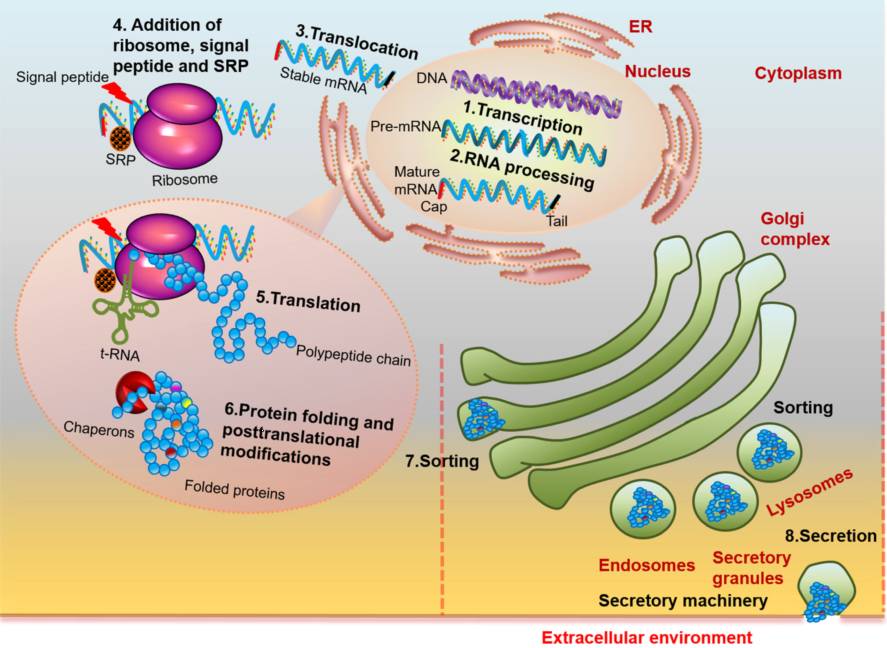
图1.抗体合成和分泌过程中所需的细胞器及过程示意图
遗传信息细胞核内由DNA转录成mRNA。mRNA前体加了5’端的帽子和3’端的polyA和剪除内含子后形成成熟mRNA,并转移至细胞质。在细胞质中,mRNA与核糖体、信号肽、信号识别粒子(SRP)结合,在tRNA的帮助下翻译成蛋白质。新和成的蛋白质经历翻译后修饰,并获得了可将其定位/分选到其正确位置的信号序列。分选后,蛋白质通过高尔基体,被包含在分泌囊泡中,分泌囊泡与细胞膜融合,将蛋白分泌至细胞外。
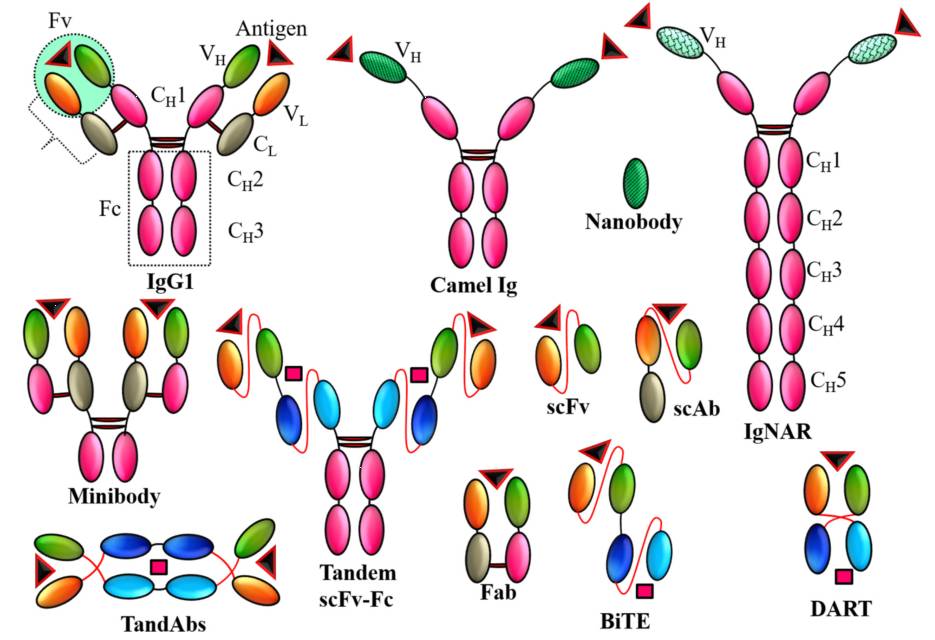
图2.抗体是由效应B细胞对抗危及宿主免疫系统的外来实体分泌的球状蛋白质
抗体分为5类:IgG、IgA、IgM、IgD和IgE,其中IgG最为普遍。抗体由两条25 kDa的轻链和两条50 kDa的重链组成。轻链和重链都分别含有恒定区和可变区。重链含有3个恒定区(C
H
1、C
H
2、C
H
3)一个可变区(V
H
),而轻链仅含有一个恒定区(C
L
)和一个可变区(V
L
)。轻链和重链并两重链间均是通过二硫键结合,形成一个Y形结构。抗体的C末端(Fc)是效应功能所必需的,而两个N末端(Fab)是抗原结合区。在整个Fab中,只有V
L
和V
H
赋予抗体靶向特异性。这认知导致许多抗体模式的发展,从简单的
SC
Fv到多价、多特异性设计的抗体模型。
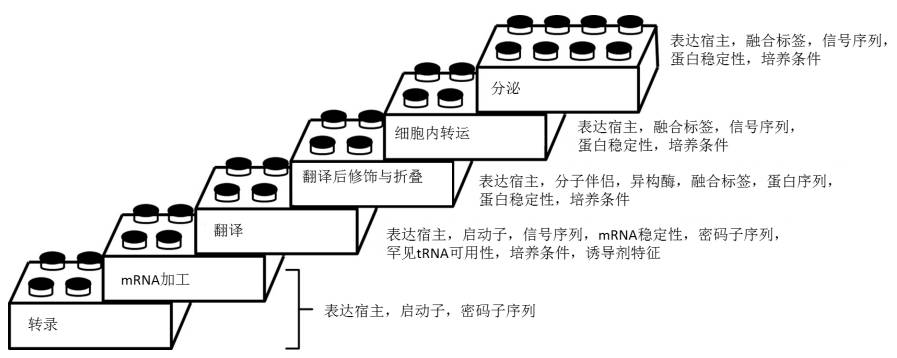
图3.建立一个有效的蛋白表达体系所包含的步骤示意图
蛋白合成包含了从转录、mRNA加工、翻译、翻译后修饰到分泌等许多步骤。此示意图概述了为获得最优的表达量,在每一个步骤中需要特别考虑的因素。
选择合适的宿主是获得最佳抗体表达的第一步。一系列的原核或者真核的宿主适用于抗体表达,它们的优劣势均列在
表1
中。哺乳动物细胞(最常见的为CHO、HEK293)是治疗性抗体的首选宿主,因为它们具有低免疫原性且其细胞器可提供最优的折叠、分泌和翻译后修饰。然而,哺乳宿主的劣势比如库体积小、生产成本高、基因操作难等,使得微生物宿主成为重组抗体生产最方便的选择。
在微生物表达宿主中,大肠杆菌无疑仍然是研究环境中被最多利用的表达系统。其受欢迎程度是由于其便利的生产和操作、便宜的培养方法、可利用的一系列载体和菌株、良好的筛选平台、快速的生产周期、无特殊要求的培养基以及简单和充分被研究的遗传信息。然而,由于氧化氛围和合适的折叠装置的限制,大肠杆菌生产具有复杂结构和富含二硫键的重组抗体的能力是有问题的。除大肠杆菌外,据报道,假单胞菌和假金丝桃属假单胞菌用于重组抗体生产,比大肠杆菌中表达量高2.5-4倍。受空间和细胞器的限制,革兰氏阴性菌分泌蛋白到周质。使用革兰氏阳性菌表达重组抗体可克服此限制,因为革兰氏阳性菌缺乏外膜,
使得蛋白可分泌至胞外培养基中。许多革兰氏阳性菌宿主,如芽孢杆菌、枯草芽孢杆菌、副干酪乳杆菌、短梗杆菌和双歧杆菌等,被开发并高产量表达分泌抗体。
酵母是另一个普遍使用的微生物表达宿主。酵母结合了原核表达系统的优势(生产周期短、成本低、基因操作简单)和真核表达系统的优势(正确折叠的翻译后修饰、分泌机制),使得它成为最受欢迎的可表达全长IgG或者其他抗体形式的宿主。
还有其他一些宿主可用于表达重组抗体,比如丝状真菌、藻类、原虫、昆虫细胞、转基因植物和动物,其中对转基因植物和动物的研究处于相对初始的阶段,需要进一步的发展。利用原核或真核转录和翻译机制的无细胞表达系统是小规模高通量快速表达功能性抗体的其他选择。 然而,这些系统对大规模表达的功效仍然是未知数。
选择宿主不仅影响表达,而且影响抗体筛选,选择和最终应用。
表1不同表达宿主的比较
|
特征
|
细菌
|
酵母
|
哺乳动物细胞
|
昆虫细胞
|
转基因植物
|
转基因动物
|
|
维护成本
|
低
|
低
|
高
|
高
|
低
|
高
|
|
载体复杂性
|
低
|
低
|
高
|
高
|
--
|
--
|
|
生产成本
|
中等
|
中等
|
高
|
高
|
低
|
高
|
|
操作难易
|
易
|
易
|
难
|
难
|
?
|
?
|
|
所需时间
|
低
|
低
|
中等
|
中等
|
高
|
很高
|
|
生长速度
|
快
|
快
|
慢
|
慢
|
慢
|
慢
|
|
载体/
菌株可获得性
|
高
|
限制
|
限制
|
限制
|
限制
|
限制
|
|
基因操纵
|
易
|
中等
|
难
|
难
|
难
|
难
|
|
发大
|
很容易
|
很容易
|
中等
|
中等
|
很容易
|
未知
|
|
繁殖
|
易
|
易
|
难
|
难
|
易
|
?
|
|
表达产品大小
|
小
|
大
|
大
|
大
|
大
|
大
|
|
二硫键
|
限制
|
无限制
|
无限制
|
无限制
|
无限制
|
无限制
|
|
产量
|
中
|
高
|
中到高
|
中到高
|
高
|
高
|
|
折叠
|
通常需重折叠
|
可能需
重折叠
|
正确折叠
|
正确折叠
|
正确折叠
|
正确折叠
|
|
糖基化
|
无
|
有
|
有
|
有
|
有
|
有
|
|
翻译后修饰
|
无
|
有
|
有
|
有
|
有
|
有
|
|
分泌
|
周质(革兰氏阴性)
胞外培养基(革兰氏阳性)
|
胞外
培养基
|
胞外
培养基
|
胞外
培养基
|
植物产物(如种子,叶子)
|
动物产物(如奶,蛋)
|
|
安全性
|
低
|
中等
|
中等
|
中等
|
高
|
高
|
丰富的表达宿主遗传学知识与优良的基因编辑策略相结合,如锌指核酸酶,转录激活子样效应核酸酶和/或CRISPR-CAS技术,使得科学家有很多机会可以系统地探索菌株修饰对抗体表达的影响。为了满足重组抗体的日益增长的需求,利用基因工程技术开发了能够表达高产量抗体的细菌、真菌、酵母和哺乳动物细胞系统(表2)。然而,一个能用于改善重组抗体表达的有效菌株的开发只能通过对影响重组抗体分泌的各种因素的进一步研究以及优化这些因素以能够产生稳定蛋白(如抗体)来实现。
表2 为改进抗体表达的宿主菌株/细胞株的修饰
|
菌株
|
修饰
|
结果
|
|
大肠杆菌
|
编码scFv
的截短基因作为包涵体过表达
|
表达的抗体纯度可达95%
|
|
大肠杆菌
|
破坏与编码两种还原酶的pET-32b
质粒序列的
N
末端融合的
trxB
和
gor
基因
|
在大肠杆菌细胞质中形成二硫键
|
|
大肠杆菌
|
共同表达
Skp
和
scFv
|
生产可溶性蛋白
|
|
酿酒酵母
|
两个ER
分子伴侣过量表达
|
增加
scFv
的表达量至
20 mg/L
|
|
米曲霉
|
重链可变区与Taka
淀粉酶
A
信号序列(
sTAA
)和来自根霉脂肪酶的
N
末端区域的
28
个氨基酸的片段的融合
|
提高λ
可变重链抗体片段的产量
|
|
泡盛曲霉
|
载体蛋白葡糖淀粉酶的利用
|
在培养基中,scFv
产量可达
50 mg/L
|
|
巴斯德毕赤酵母
|
抑制N-
链接糖基化的作用的糖基化工程
|
分泌的重组蛋白具有高度均一的N-
连接聚糖
|
|
Ogataea minuta
|
抑制液泡蛋白酶和天冬氨酸蛋白酶对分泌的抗体的作用
|
抑制了分泌的抗体的部分降解
|
|
利什曼原虫
|
分泌信号肽与scFv N
末端连接
|
观察到不同的翻译后修饰能力
|
|
烟草本鼻烟
|
pE16
变体由一个与人
IgG
的重链恒定结构域融合的
scFv
组成
|
糖分析显示哺乳动物糖基化谱
|
|
小立碗藓
|
移除植物特异性N-
聚糖结构
|
分泌的抗体ADCC
作用增强了
40
倍
|
|
中国仓鼠卵巢细胞(CHO
)
|
CHOP
单独或与
URP
相关基因如
CHOP / Gadd153
和
GRP78
或
ER
伴侣蛋白的超表达
|
鉴定具有高产量的功能性抗体的细胞系
|
丰富的表达宿主遗传学知识与优良的基因编辑策略相结合,如锌指核酸酶,转录激活子样效应核酸酶和/或CRISPR-CAS技术,使得科学家有很多机会可以系统地探索菌株修饰对抗体表达的影响。为了满足重组抗体的日益增长的需求,利用基因工程技术开发了能够表达高产量抗体的细菌、真菌、酵母和哺乳动物细胞系统(表2)。然而,一个能用于改善重组抗体表达的有效菌株的开发只能通过对影响重组抗体分泌的各种因素的进一步研究以及优化这些因素以能够产生稳定蛋白(如抗体)来实现。
表2 为改进抗体表达的宿主菌株/细胞株的修饰
|
菌株
|
修饰
|
结果
|
|
大肠杆菌
|
编码scFv的截短基因作为包涵体过表达
|
表达的抗体纯度可达95%
|
|
大肠杆菌
|
破坏与编码两种还原酶的pET-32b质粒序列的N末端融合的trxB和gor基因
|
在大肠杆菌细胞质中形成二硫键
|
|
大肠杆菌
|
共同表达Skp和
scFv
|
生产可溶性蛋白
|
|
酿酒酵母
|
两个ER分子伴侣过量表达
|
增加
scFv的表达量至20 mg/L
|
|
米曲霉
|
重链可变区与Taka淀粉酶A信号序列(sTAA)和来自根霉脂肪酶的N末端区域的28个氨基酸的片段的融合
|
提高λ可变重链抗体片段的产量
|
|
泡盛曲霉
|
载体蛋白葡糖淀粉酶的利用
|
在培养基中,scFv产量可达50 mg/L
|
|
巴斯德毕赤酵母
|
抑制N-链接糖基化的作用的糖基化工程
|
分泌的重组蛋白具有高度均一的N-连接聚糖
|
|
Ogataea minuta
|
抑制液泡蛋白酶和天冬氨酸蛋白酶对分泌的抗体的作用
|
抑制了分泌的抗体的部分降解
|
|
利什曼原虫
|
分泌信号肽与scFv N末端连接
|
观察到不同的翻译后修饰能力
|
|
烟草本鼻烟
|
pE16变体由一个与人IgG的重链恒定结构域融合的scFv组成
|
糖分析显示哺乳动物糖基化谱
|
|
小立碗藓
|
移除植物特异性N-聚糖结构
|
分泌的抗体ADCC作用增强了40倍
|
|
中国仓鼠卵巢细胞(CHO)
|
CHOP单独或与URP相关基因如CHOP / Gadd153和GRP78或ER伴侣蛋白的超表达
|
鉴定具有高产量的功能性抗体的细胞系
|
基因表达是一个涉及多个调节因子的受控过程,并在转录和翻译水平上影响整个表达过程。一些因素可以影响基因表达水平,如每个氨基酸的密码子使用频率,mRNA 50’末端折叠成稳定的二级结构的趋势,mRNA的GC含量,以及转录终止子的存在,RNA降解结构基元和DNA重组位点(如图1所示)。遗传密码的退化导致氨基酸(甲硫氨酸和色氨酸除外)由多个(2-6)同义密码子编码。 在给定的宿主中,不是所有的密码子都以相同的频率使用;有些被广泛用于高度表达的基因(最佳密码子),而有些则很少被使用(稀有的密码子),这又反映了它们的同源tRNA的丰度。当靶基因的最佳密码子的使用频率与表达宿主显著不同时,密码子偏倚现象就会出现。密码子偏倚对表达量的影响程度仍然具有争议,但很明显,密码子偏倚会影响翻译的几个方面,包括氨基酸插入的准确性,多肽折叠,mRNA序列和稳定性。密码子偏倚可以通过靶基因的密码子优化来解决,以适应宿主的tRNA库,或通过修饰宿主以增加代表性不足的tRNA的可用性。
抗体中的可变区包含来自于VDJ重组和体细胞高频突变的高度不同的序列,使得抗体具有特异性和亲和力。这些不同的序列有时会损害重组抗体的稳定性,使其易于聚集并降低表达量。针对改善/增强抗体生物物理性质的持续努力已确定一些影响重组抗体稳定性的内在和外在因素,如表面暴露残基,可变区界面的疏水核心和最佳V
H
-V
L
配对。Demarest报道了通过在Fv区域中突变11个残基,可增加Fab的稳定性和功能性表达。 另一项研究表明,另一项研究表明,用带负电荷的氨基酸(天冬氨酸或谷氨酸)替代高度特异性位置的氨基酸(V
H
:28, 30, 31, 32, 33, 35; V
L
: 24, 49, 50, 51, 52, 53, 56)不仅可以改善表达,还能使抗体无聚合。有时,V
H
和V
L
结构域在文库构建阶段时的随机组合会导致在体内发生和选择的天然配对的丧失,并需要额外的亲和力成熟步骤来优化所得重组抗体的亲和力和稳定性。这表明V
H
和V
L
相互作用在确定重组抗体的生物物理性质方面的重要性。糖基化也可以通过改变其稳定性来影响重组抗体的表达。抗体在每个重链的CH2 Asn297上具有N-连接的糖基化位点,而糖基化在抗体效应子功能和药代动力学中起重要作用。不同的表达宿主的翻译后修饰机制各不相同,这很大程度地影响它们所表达的抗体的糖基化修饰情况。
抗体的形式可以作为抗体表达的决定因素。结构域顺序、接头性质和标签存在均可影响重组抗体的表达,特别是scFv的表达。在一个scFv的设计中,可以是V
L
-V
H
或者V
H
-V
L
,一个结构域的3’端连着另一个的5’端;有研究显示,可变区中结构域的顺序以及接头的长度会显著影响scFv的产量,甚至可达100倍。接头是重组抗体的关键组分,它可以影响抗体的灵活性、稳定性和抗原结合特性。接头的长度和氨基酸种类均可影响抗体的热稳定性和表达。通常,使用含有甘氨酸和丝氨酸残基组合的接头会所得重组抗体具有灵活性和蛋白酶抗性。然而,这样的组合并不能保证对所有表达的抗体来说都是最佳选择。通过增加一个融合蛋白也是改进抗体的产量、增强其稳定性和溶解度特性的普遍方式。










