编码和非编码RNA的转录后修饰在细胞中是普遍存在的。
N6-甲基腺苷(m6A)是真核信使RNA(mRNAs)上最丰富的修饰类型.这种修饰在多种基本生物过程中起着重要作用,例如细胞分化,组织发育和肿瘤发生。
像许多其他生物大分子的化学修饰,也可以被m6A特定识别。
2019年8月6日,清华大学李丕龙等人在
Cell Research
杂志在线发表了题为“
Multivalent m6A motifs promote phase separation of YTHDF proteins
”的研究论文,该研究发现
含多价m6A的RNA可以增强体外和细胞中YTHDF蛋白的相分离潜力。
细胞生物化学的一个重要组成部分是非膜性区室中蛋白质和核酸的浓度。这些生物分子缩合物由包括液 - 液相分离的过程形成。
液 - 液相分离所需的多价相互作用已在体外进行了广泛研究。
然而,人们对体内该过程的调节知之甚少。2019年5月1号,清华大学李丕龙研究团队等人在
Nature
在线发表了题为“
Arabidopsis FLL2 promotes liquid–liquid phase separation of polyadenylation complexes
”的研究论文,该研究结果表明,
卷曲螺旋域蛋白可以促进液 - 液相分离,这扩展了我们对控制液体样体内动力学原理的理解。

N6-甲基腺苷(m6A)被认为是在真核生物中最普遍的信使RNA(mRNA)内部修饰,包括在哺乳动物,植物,果蝇和酵母中都存在
。m6A修饰在mRNA代谢中发挥多种功能,包括mRNA稳定性,剪接,翻译效率和核输出。越来越多的证据表明,m6A影响不同的发育和生物过程,如癌症干细胞增殖,胚胎和胚胎发育后期,细胞昼夜节律和细胞命运决定等,突出了m6A修饰生物学的重要性。
作为一种动态和可逆的转录后修饰,哺乳动物中的m6A甲基化通过甲基转移酶复合物获取,该复合物含有甲基转移酶如3(METTL3),METTL14和Wilms'肿瘤1相关蛋白(WTAP),它的去除是由去甲基化酶FTO和烷基化DNA修复蛋白AlkB同系物5(ALKBH5)介导的。
通过“读取”蛋白(例如YTH结构域家族蛋白)实现对m6A修饰的转录物的识别,其介导m6A修饰的下游效应。
文章总结
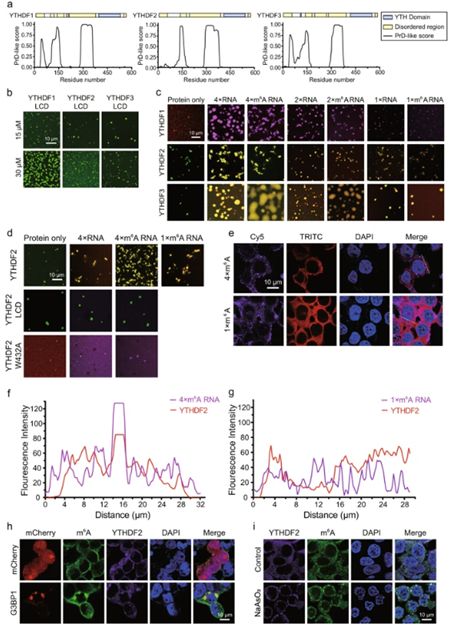
研究最深入的m6A读取子结构域是YT521-B同源(YTH)结构域,其从酵母到人类保守并且优先结合RR(m6A)CU(R = G或A)共有基序。
人类有五种含有YTH结构域的蛋白质,其中三种YTHDF1-3属于同一蛋白质家族。除了C末端折叠良好的YTH结构域外,所有三种蛋白质也是如此,
在N端具有低复杂度域(LCD):
预计它们的LCD区域是Prion样区域。
含有朊病毒样结构域的蛋白质通常具有相分离的可能性。
通常发现相分离有助于生物分子缩合。事实上,所有三种YTHDF蛋白质的LCD都经历了浓度依赖性相分离。
尽管与它们的LCD相比相分离能力降低了,但是全长YTHDF1 / 2/3蛋白在没有RNA的情况下也在生理条件下进行相分离。
在这项研究中,研究人员发现含
多价m6A的RNA可以增强体外和细胞中YTHDF蛋白的相分离潜力。
这些数据表明m6A和YTHDF2的相分离与细胞对应激的反应有关。
m6A修饰的功能是多种多样的,并且其中许多是通过其读取蛋白识别m6A来进行的。
对于m6A的许多上述功能,YTHDF的多价m6A驱动的相分离可能是重要的。
参考信息:
https://www.nature.com/articles/s41422-019-0210-3

—
END
—
本
文仅限转载,不代表本公众号观点