编者按:
在很多人看来,生命体的遗传信息流,由DNA流向RNA,再通过翻译,流向蛋白质。中心法则中,RNA的地位有些尴尬,既不是遗传信息的储存者,又不是生命活动的主要执行者,似乎就是一个两头跑的打工仔,在DNA和蛋白质面前却要矮上三分。实际上,RNA很可能是一位隐藏很深的大Boss,没有它,DNA的政令不出细胞核,甚至不出染色质。本专栏由复旦大学教授于文强组织策划,邀请了国内外表观遗传学领域工作者共同完成。了解表观遗传领域近年的发展概况以及解释我们日常有意思的生命现象是我们开设此专栏的初衷。
撰文 | 段洪超(北京大学化学与分子工程学院博士生)
责编 | 叶水送
● ● ●
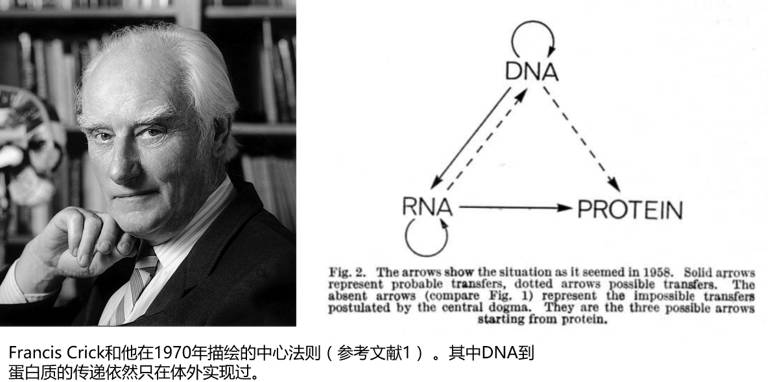
英国著名生物学家弗朗西斯·克里克提出中心法则的时候,一定想不到他的后辈们在上面玩出这么多新花样。绝大部分生命体会把遗传信息储存在DNA中,遗传信息通过转录流向RNA,再通过翻译流向蛋白质,蛋白质是生命活动的主要执行者。在RNA病毒中,遗传信息又会有RNA到DNA和RNA到RNA两个流向,不过这并不是生命历史发展的主要进程。
随着一代代生物学家的不懈努力,人们发现遗传信息的流动还被一些额外的化学修饰所操控,比如DNA的甲基化、羟甲基化,组蛋白的甲基化、乙酰化等等,这些修饰导致在DNA序列没发生改变的情况下,基因的表达却改变了,这就是所谓的表观遗传修饰。
我们可能隐隐约约地有种感觉,就是在原版的中心法则中,RNA的地位有些尴尬,它既不是遗传信息的储存者,又不是生命活动的主要执行者,似乎就是一个两头跑的打工仔,虽然没它不行,可在DNA和蛋白质面前却要矮上三分。实际上,RNA很可能是一位隐藏很深的大Boss,它的工作出了一点偏差,分分钟就让DNA政令不出细胞核——甚至政令不出染色质。
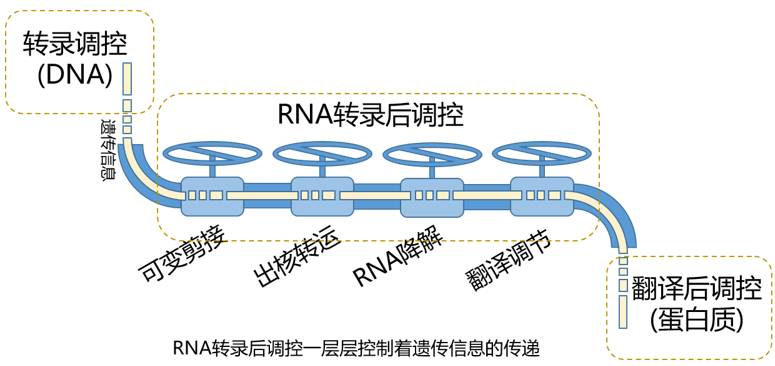
事情是这样的。外面世界太险恶,生命体必须演化出一套复杂的表达调控机制,涉及DNA、RNA和蛋白质各个层面。把它类比到一个生活化的场景中,针对DNA的转录调控就如同种粮食,这是一个最基本的环节,但是由转录得到的产物就好比刚从地里收获的庄稼,还需要经过严格的质控,精细的加工,高效的分销,变成大米面粉,或者馒头面包才能进入各家各户,这就类似于RNA转录后调控的作用。外部环境的变化通常是快速而剧烈的,DNA水平的转录调控往往反应不够迅速,或者在精细程度上相对缺失,这就需要RNA水平的转录后调控出来独当一面。
由此产生的结果便是,同样的转录水平,产生了不同数量的蛋白质,甚至同样的基因序列,制造出不同序列的蛋白质。RNA的可变剪接、转运出核、降解以及翻译水平调节,这些复杂的转录后调控过程,精准操纵着遗传信息的流动,使生命体从容应对千变万化的外部环境。
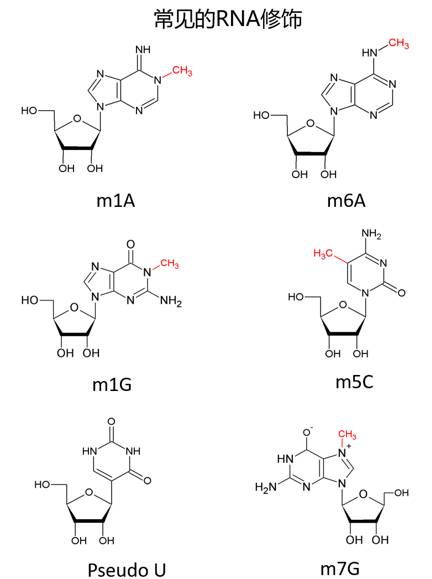
与DNA相同,RNA也把最基本的遗传信息承载于四种碱基上——只不过是胸腺嘧啶T换成了尿嘧啶U——那它是如何完成了DNA无法做到的任务呢,一部分是通过各种或长或短的非编码RNA来起作用,但更主要的是通过丰富的RNA修饰来完成。——事实上,很多非编码RNA上的修饰也极大地影响了它们的功能。修饰就好比穿马甲,穿上马甲之后你就换了一个角色,隔壁的那个蛋白质就不认得你了。DNA的马甲就比较少,在绝大部分高等生物的DNA上,除却5-甲基胞嘧啶(5mC)和5-羟甲基胞嘧啶(5hmC),其它的修饰极少见。RNA的任务比较多,所以RNA就有好多马甲。迄今为止,人们在RNA上已经发现了一百多种不同的修饰,而且其中很多修饰在进化上都十分保守。试想一下,你我都有一件十几亿年前就存在于地球上的马甲,也是件令人激动的事情。
穿上马甲的RNA是怎样扮演不同角色的呢?由于生命活动的主要执行者是蛋白质,马甲也主要是通过影响特定的结合蛋白来完成的。这些结合蛋白大致可分为两类,一类是识别特定RNA修饰的蛋白,这叫做“非你不可”型的,另一类是识别特定的RNA结构——RNA会通过碱基间的氢键和π-π相互作用修饰形成高度复杂有序的结构,而RNA修饰会导致这种结构的改变——这叫做“爱屋及乌”型的。这样,同一位点不同程度的修饰便会与不同水平的结合蛋白相结合,而这些结合蛋白将会招募相应的分子机器,完成RNA的可变剪接、转运出核、降解以及翻译水平调节等过程。穿上了不同的马甲,就招呼来了不同的小伙伴,而正是这些小伙伴赋予了RNA修饰真正的意义。这也就使得对RNA修饰结合蛋白的研究成为这一领域中决定性的部分。
通过RNA修饰介导的转录后调控,基因的表达在其序列没有发生改变的情况下出现了差异,这使得RNA表观遗传学(RNA epigenetics)这一很先锋的概念逐渐成型。有些人不太想搞大新闻,他们更倾向于接受Epitranscriptomics(表观转录组学)这一温和的说法,但不可否认的是,RNA修饰的研究正与传统表观遗传学研究进行着越来越深入的互动,共同推动着生命科学领域的一场风暴。
2007年,不堪身体之重的欧洲人终于发表了第一个肥胖相关的SNP(单核苷酸多态性),意外的是,这个SNP并不位于诸如瘦素或者瘦素受体等早已被克隆的肥胖相关基因,而是位于名不见经传的基因KIA1005上。这一基因随后被命名为FTO(fat mass and obesity associated)。而对FTO分子功能的研究发现,它在体内的底物是N6-甲基腺嘌呤(m6A,这个马甲在腺嘌呤的身上写了个大大的M)这一mRNA上最为丰富的修饰——它可以通过氧化去甲基作用,让RNA把穿上的马甲脱下来。
早先人们通常认为,当RNA修饰完成时,这一条RNA的命运便就此决定了,它将沿着确定的路线发挥功能直到降解。然而现在人们知道,至少有一种RNA修饰是可逆的。这表明RNA手里的并非拿着单程票,而是往返票,它脱下马甲还可以行使原来的角色。无巧不成书,其实就在那一年,研究者第一次确认,哺乳动物DNA上的5-甲基胞嘧啶修饰也是可逆的,成为与这一最重要表观遗传学标记交相辉映的双子星,人们对RNA修饰,特别是m6A修饰的研究热情突然间高涨了起来。
对RNA修饰的研究大体分为三个部分,第一是修饰酶,通常把它叫做Writer,就是向RNA引入这一修饰的家伙;第二是脱修饰酶,就是脱马甲的那个家伙,通常把它叫做Eraser,它将RNA从被修饰状态逆转为非修饰状态;第三是结合蛋白,通常把它叫做Reader,前边已经讲到,对RNA修饰结合蛋白的研究,是这一领域中决定性的部分,而对结合蛋白的研究,也往往能为通过RNA修饰解决具体的生物学问题,提供一个模型或范式。m6A第一个被确认的结合蛋白是YTHDF2,YTHDF2会介导其底物mRNA的降解,作为YTHDF2的底物,不含m6A修饰的mRNA会比含有这一修饰的mRNA更稳定。换句话讲,m6A修饰关系到mRNA是“怎么没的”。通常地,基于DNA的转录调控只能告诉我们RNA是怎么来的,可如果它该没的时候还在,该在的时候却没了,就一定会出问题。
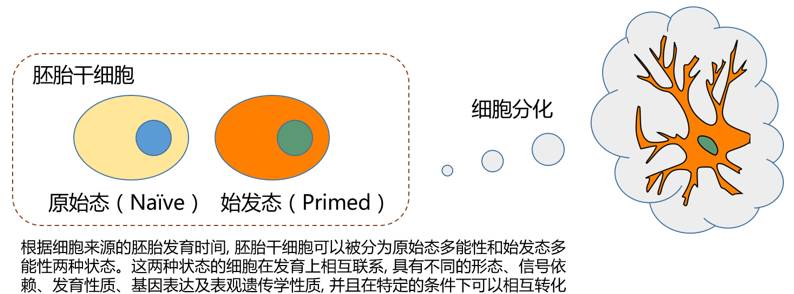
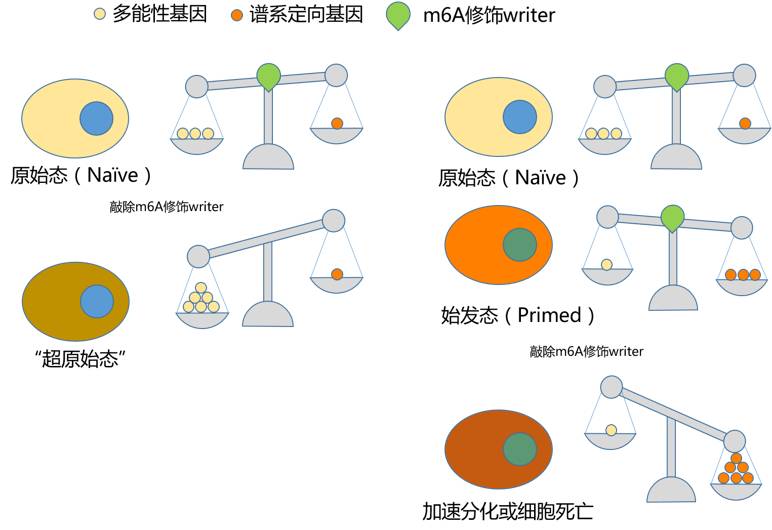
干细胞分化过程就是一个这样的例子。在胚胎干细胞中,分化或不分化,主要靠两类基因说了算,一类是使其保持原始态多能性的基因,另一类是使其谱系定向调控基因。两类基因就如同天平的两侧,哪一侧基因的表达量占优,干细胞就会倾向于哪一侧的状态。两类基因都能抑制对方的表达,干细胞就是通过这种方式,维持状态的持续性和稳定性。而诸多原始态多能性保持和谱系定向调控基因的mRNA都存在m6A修饰。这使得相关基因的mRNA处在一种“说没就没”的状态。在原始态多能性状态下,让干细胞不分化的基因处于优势位置,m6A修饰的缺失使这些基因mRNA的稳定性提高,赖着不走,又对谱系定向调控基因施加了持续的抑制作用,从而导致干细胞进入一种“超原始”的状态,阻止其分化。而在始发态多能性状态下,让干细胞分化的基因表达开始上升,m6A修饰的缺失使这些基因mRNA的稳定性提高,最终导致干细胞加速分化或细胞死亡。
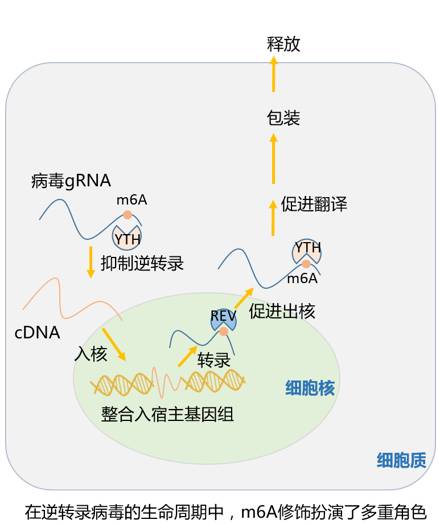
不仅仅是生物内源的RNA,像RNA病毒这种外源的入侵者也存在RNA修饰。很早之前人们就已经在诸如流感病毒、肉瘤病毒这样的RNA病毒上发现了m6A修饰。不过其对病毒本身的影响最近才逐渐为人所知,而且有一点众说纷纭,莫衷一是的感觉。总的来说,对于HIV、流感病毒这样的逆转录病毒,由于不同结合蛋白的不同功能,在其没整合进基因组之前,m6A的存在对它们有抑制作用,但如果它已经整合进基因组,m6A就要老老实实帮病毒做事了。
对于寨卡病毒和丙肝病毒这样的正义单链RNA病毒,m6A在其生命周期中起到了抑制的作用。乍看起来m6A在帮着宿主虐病毒,然而测序结果显示,丙肝病毒的m6A修饰位点在进化中体现出较高的保守性,人家十万八千辈儿都穿着这个马甲。由于病毒通常会面对很强的选择压力,保守的m6A修饰位点似乎又暗示其在丙肝病毒生命周期中的积极作用。总而言之,m6A修饰在RNA病毒中的研究方兴未艾,由于许多RNA病毒均含有m6A修饰,以此通路为靶点有望开发出可抗击多种病毒病的药物。
就好比织出的布不能直接穿在身上,而要经过剪裁和缝纫才能变成衣服,由DNA直接产生的RNA半成品中,很多是不会包含在成熟RNA中的,它们要在核内由相应的机制去除,这就是RNA的剪接。而正如不同的人有着不同的体型、衣服的尺码也会不同,RNA在不同的情况下,也会有不同的剪接方式。m6A修饰在这边的角色,就如同裁缝手里的一把尺子,指示了RNA剪接发生的位置。它首先被结合蛋白所识别,随后m6A结合蛋白会继续召唤从事RNA剪接的小伙伴,所以剪接发生的位点通常在距m6A修饰不远的地方。
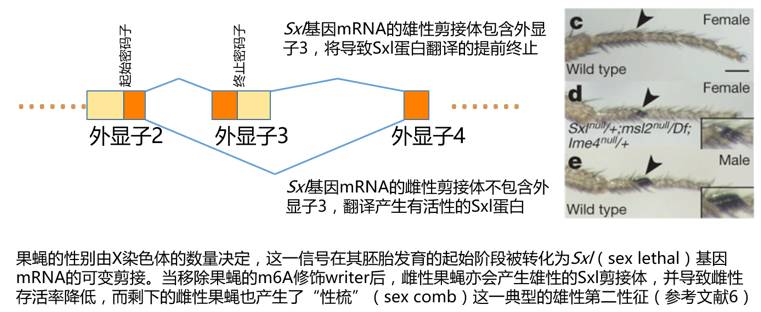
RNA的可变剪接影响了众多生命过程,其中最脍炙人口的要数果蝇的性别决定了。果蝇的性染色体也是XY型(雄性XY,雌性XX),与和哺乳动物不同的是,哺乳动物性别看有无Y染色体,而果蝇则要看有多少X染色体,带够了两条X染色体为雌果蝇,只有一条X染色体就只能是雄果蝇了。通过果蝇的繁殖实验研究者发现,X染色体数量信号,影响了一个叫Sxl(Sex lethal)基因mRNA的可变剪接。在雌果蝇当中,Sxl基因的mRNA会被正常剪接,并产生正常的Sxl蛋白,而在雄果蝇中,Sxl基因的mRNA剪接异常,导致雄果蝇只能产生截短的Sxl蛋白。Sxl蛋白的不同会带来一系列基因的差异表达,并最终决定雌雄果蝇的性征。而在Sxl的mRNA前体中,可变剪接位置附近就包含了m6A修饰。当m6A修饰的Writer被移除之后(这样可变剪接位置附近就没有m6A了),雌果蝇中出现了像雄果蝇一样的Sxl剪接体,而雌果蝇的存活率也大幅下降了,存活下来的雌果蝇很多也出现了雄果蝇的性征。总结一下,如果没有m6A修饰,好多雄果蝇就会当光棍,剩下的许多雌果蝇也长出“大胡子”。
RNA修饰是一个内容极其丰富的研究领域,限于篇幅只能撷取极为有限的内容进行介绍。事实上许多重要的RNA修饰,比如假尿嘧啶修饰(pseudo U)和N1-甲基腺嘌呤(m1A),都有着丰富而精彩的故事值得讲述,它们对生物响应外界刺激,维持自身内部稳态,都有着不可替代的作用。自上世纪六七十年代发展至今,这一领域已发展出众多独到的研究方法,也积累了众多重要的科学事实。
近年来,借由高通量测序技术的进步以及与其它学科交叉的成果,RNA修饰领域逐渐连点成线,连线成面,开始向世人展示出一副极其绚烂的科学画卷,尽管其研究的深度在目前还不及DNA和染色质的表观遗传修饰,但其研究的广度却是后者远不能及的,相信在不久的未来RNA在中心法则的核心位置将愈发显现出来。该领域最新的研究结果,逐步揭示RNA修饰与流感、艾滋病、白血病、阿尔茨海默病密切的联系。无独有偶,在高等植物中,RNA修饰亦影响着其胚胎发育、开花结果,乃至于抗病、抗非生物胁迫的生命过程。这都预示着RNA修饰的研究必将深刻地影响医疗、制药、乃至农业的发展。
表观遗传学专栏:
20170112 生命之初,新在哪里:我们为何生而不同
20170121 父母基因博弈,谁赢你像谁?
20170214 曾与诺奖失之交臂、人小鬼大的miRNA如何改变细胞命运?
20170303 人类克隆自己:还有什么困难?
制版编辑:叶水送丨
本页刊发内容未经书面许可禁止转载及使用
公众号、报刊等转载请联系授权
[email protected]
▼点击查看相关文章
该不该学奥数|数学巨匠|五音不全|抗癌药
新年献词|最受欢迎|西湖|农场|学术辩|日本奖
屠呦呦|王晓东|白岩松|何江|张锋|杨振宁|李佩
卢煜明|王小凡|霾|女性成就|张纯如|数学教皇

▼▼▼点击“阅读原文”,牵手科学队长!










