指唾液酸在
糖蛋白末端的共价加成
,参与胚胎发育、神经发育、重编程、肿瘤发生和免疫反应过程。
唾液酸化
是指
唾液酸基团的末端添加到寡糖和糖蛋白的末端
。
唾液酸
是一个由九碳酮基酸性单糖及其衍生物组成的大家族,通常
附着在聚糖的远端
,使它们成为细胞之间以及细胞和细胞外基质之间的
“桥梁”分子
。
目前已发现唾液酸由
N-乙酰神经氨基酸(Neu5Ac)
、
N-糖基神经氨基酸(Neu5Gc)
、
脱氨基神经氨基酸(Kdn)
及其衍生物组成。唾液酸通过
α-2、3-或α-2、6-键连接到O-2和N-联聚糖
,或通过
特定酶的α-2、8-或α-2、9-键
连接到其他唾液酸部分
。
唾液酸化聚糖表现出
广泛的结构多样性
,这使它们在不同的过程中具有丰富的生物学功能,包括
发育、体细胞重编程和癌症进展
。
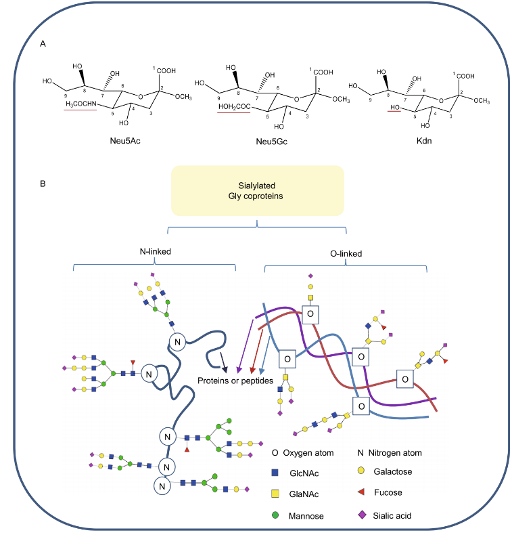
图1
唾液酸的结构和唾液酸化糖蛋白的多样性
Neu5Ac
和
Neu5Gc
是两种重要的唾液酸,它们的结构差别仅仅在于1个氧原子。
细胞质中,在胞苷单磷酸N-乙酰神经氨酸羟化酶
CMAH
催化下,Neu5Ac以
CMP-Neu5Ac
的形式被还原,即Neu5Ac中的N-乙酰基被还原成
N-羟乙酰
,CMP-Neu5Ac转化成
CMP-Neu5Gc
。
CMP-Neu5Ac和CMP-Neu5Gc
二者都可
作为唾液酸供体加到多种糖复合物上
。
Neu5Ac和Neu5Gc
存在于大多数哺乳动物组织中
。
哺乳动物的
唾液酸和唾液酸化聚糖的生物合成途径
已经被揭示出来,超过20多种
唾液酰基转移酶
已被鉴定。
Neu5Ac
是最典型的唾液酸,由
UDP-N-乙酰氨基葡萄糖(UDP-GlcNAc)
合成,进而由细胞质中的
己糖胺途径
产生。
UDPGlcNAc
首先通过
UDP-GlcNAc2-异构酶
转化为唾液酸的第一个前体(
ManNAc
),将
ManNAc
转化为
ManNAc-6P
,并
通过
N-乙酰神经胺-9-磷酸合成酶
转化为Neu5Ac-9P。
在胞质的最后一步,
N-酰基神经氨酸-9-磷酸酶
将
Neu5Ac-9P
转化为
Neu5Ac
,Neu5Ac进入细胞核,通过
CMP-NeuNAc合成酶
转化为
CMP-Neu5Ac
。一部分CMP-Neu5Ac被
CMP-Neu5Ac羟化酶
转化为
CMP-Neu5Gc
。
这些核苷酸糖被运输到
高尔基体
,在那里
产生
唾液-糖缀合物
。
最后,唾液酸糖蛋白被
水解,可以再生唾液酸和
回收来合成更多的
唾液酸-糖缀合物
。
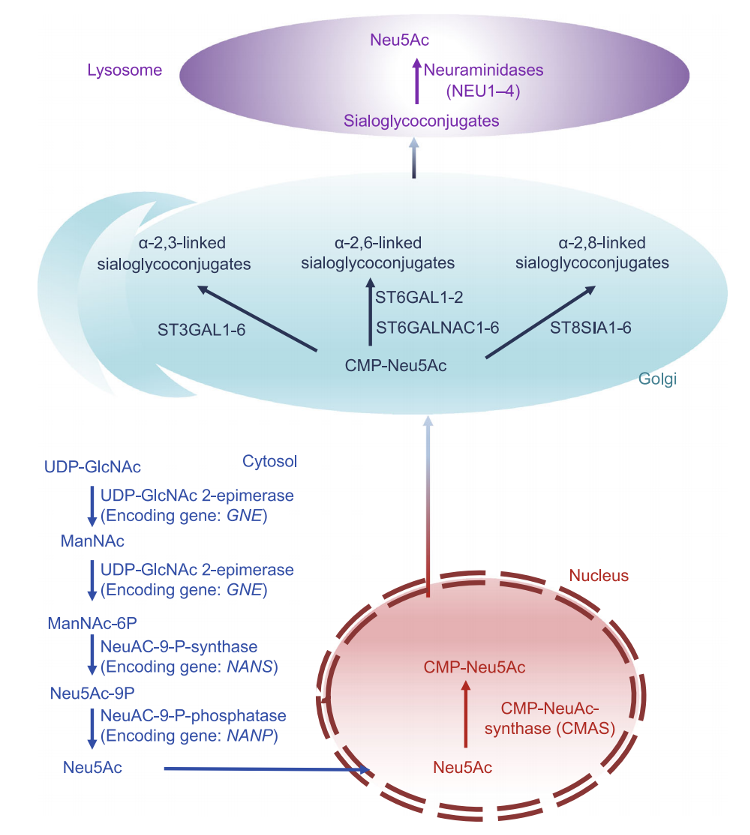
图2
唾液酸化的生物合成途径
唾液酰化
在早期胚胎发育中的作用在2002年首次报道。缺乏
UDP-GlcNAc2-外构酶
编码基因Gne等位基因的杂合子小鼠没有出现异常,但没有引起任何纯合子敲除,表明了Gne失活的
早期胚胎死亡率
。
在发育过程中,细胞间的粘附激活了对
生存、迁移和分化
至关重要的信号通路。许多参与
早期胚胎发育
的细胞粘附分子是唾液酸化糖蛋白,如E-cadherin、整合素和连环蛋白。
异常的唾液酰化
可以抑制这些粘附分子与其受体之间的相互作用,从而阻断与发育过程相关的信号转导。
Cmas
介导的
唾液酰化
在体外早期小鼠胚胎发育中是不可或缺的。该基因
缺失导致
细胞表面唾液酸化的完全丢失
。
多能干细胞
具有无限期的
自我更新和分化为身体所有细胞
的能力,这使得多能干细胞,包括
胚胎干细胞(ESCs)
和
诱导多能干细胞(iPSCs)
,对于特定研究和临床应用具有重要价值。
至今,我们仍未完全了解这些细胞在细胞重编程过程中如何
建立、维持和调节它们的多能性
。
最近研究表明,
唾液酸化对干细胞多能性的建立和维持至关重要
,
蛋白唾液酸化水平在细胞分化过程中显著改
变
。
与非多能性细胞相比,未分化的多能干细胞中
ST6GAL1唾液酰基转移酶
水平更高。此外,
ST6GAL1基因
的敲除
以及
唾液酰基转移酶抑制剂
的存在
降低了体细胞重编程的效率
。
唾液酸在生理pH时给细胞表面糖蛋白带负电荷,影响
构象和寡聚
以及与
其他细胞和细胞外基质蛋白的相互作用
。
此外,
唾液化聚糖
是控制关键生物过程的许多蛋白质的
配体
,包括
恶性转化
。
一些研究表明
正常细胞向异质癌细胞的转化伴随着
异常的蛋白唾液酸化修饰
。
因此,在癌细胞上改变的唾液酸化可以作为
潜在的生物标志物,
包括
总唾液酸、唾液酸化的糖蛋白和碳水化合物抗原
。
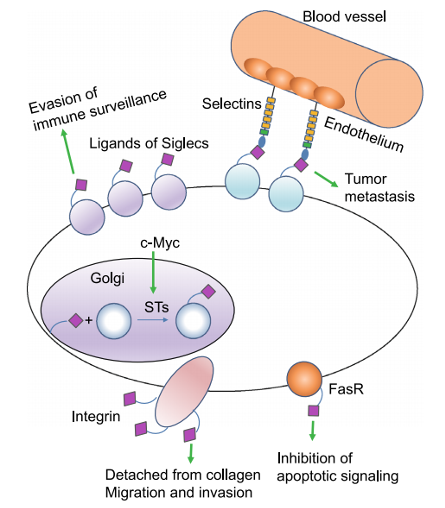
图3
癌症中的唾液酸化
如果有基金申请或者课题思路疑问,
请联系Alice咨询:






