今天给大家介绍的是发表在Nature上的“
Adipose-derived circulating miRNAs regulate gene expression in other tissues
”,即
脂肪来源的循环
miRNAs可调节其他组织的基因表达。
miRNA是一种短链非编码RNA,可作为一种阴性调节因子参与许多细胞进程。某些特定的miRNA的水平变化与许多疾病相关,如癌症、糖尿病、心血管疾病等。脂肪组织能产生大量的
miRNA。
白色脂肪组织(WAT)中的
miRNA
也随年龄增加而下降(
因为miRNA处理酶Dicer随着年龄增加的减少
),同样的情况也发生在HIV相关的脂肪代谢障碍的人群。已有研究发现在循环血液中有很多包含有miRNA的外泌体。为了研究脂肪组织中miRNA的作用,本文作者建立了脂肪组织中缺乏Dicer酶的小鼠模型(AdicerKO小鼠),然后从小鼠血液中提取了外泌体并且分析了外泌体中的miRNA,发现相对于正常小鼠,外泌体中有422种miRNA含量发生了显著的变化,其中419种显著下降(FIG1 a-c),
在
先天性全身脂质营养不良(CGL)与HIV相关营养不良患者中也发现了相似的情况(FIG1 d-g),说明脂肪组织是外泌体miRNA的主要来源。
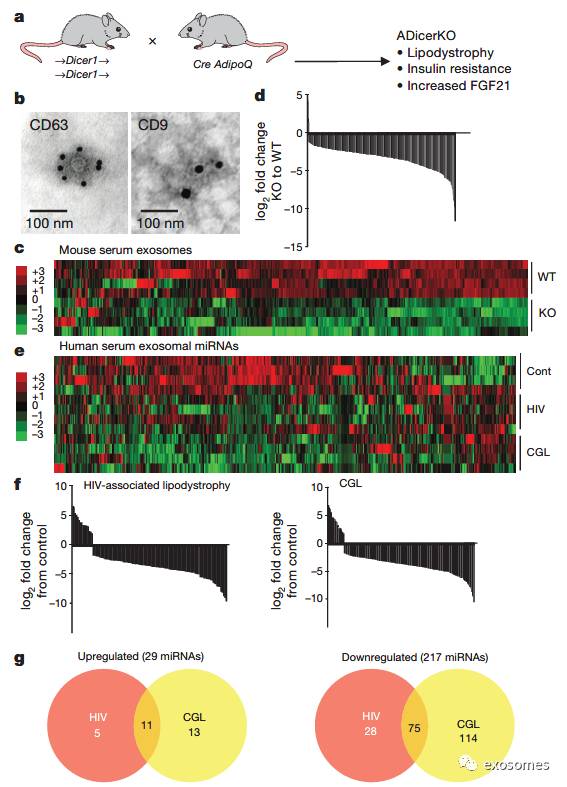
图一:外泌体miRNA主要来源于脂肪组织。
为了证明脂肪组织是外泌体miRNA的主要来源,作者将正常小鼠的腹股沟、附睾的WAT及棕色脂肪组织(BAT)移植入
AdicerKO
小鼠,发现有超过50%的miRNA表达恢复(FIG2 a-c)。并且发现AdicerKO小鼠糖耐量异常,且其糖耐量只在移植入BAT时恢复正常。(FIG2 d-e)
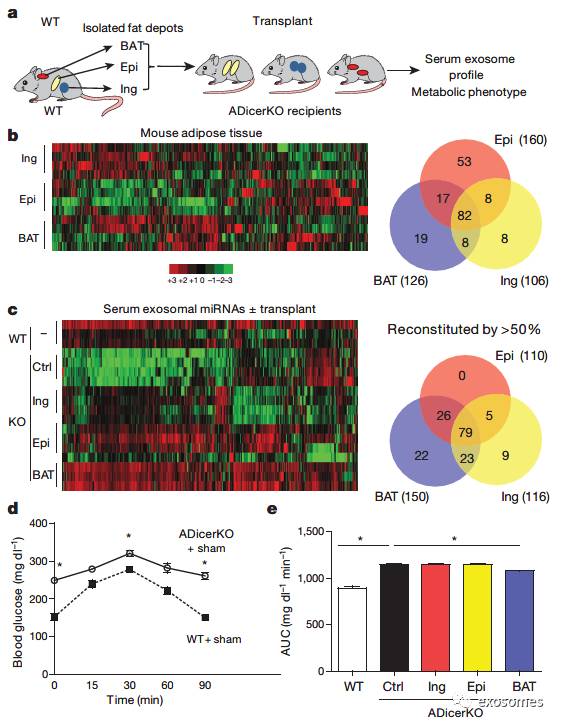
图二:脂肪移植增加
AdicerKO小鼠
的循环外泌体miRNA。
进一步研究发现,
AdicerKO
小鼠的循环中的
FGF21
(成纤维细胞生长因子,可影响多种组织的代谢)是正常小鼠的3倍,且其肝脏、胰腺、肌肉及脂肪组织中的
Fgf21 mRNA
明显增加(FIG3 a-b)。将脂肪组织移植入AdicerKO小鼠,发现只有在移植入BAT之后,血液和肝脏中的Fgf21 mRNA明显减少,这说明BAT能生成某种能直接或间接调节肝内FGF21表达的因子。假设这种因子是循环的miRNA,于是作者查找了miRDB数据库,发现了4种可能相关的miRNA,其中3种(miR-99a, miR-99b, and miR-100)在
AdicerKO小鼠中
明显减少。之后作者在外泌体中的导入这几种miRNA,并将外泌体转入小鼠体内,发现在导入了
miR-99b
之后,Fgf21 3’端活性明显减少。(FIG3 c-f)
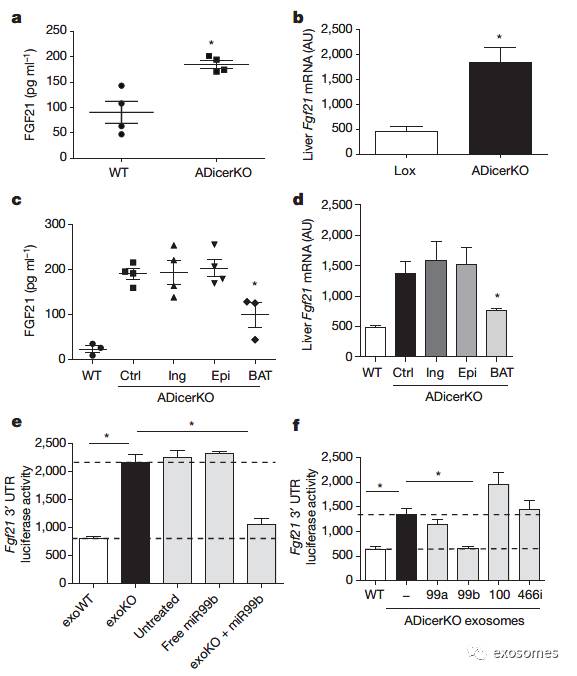
图三:脂肪组织来源的外泌体miRNA能调节肝脏FGF21及其转录。
作者将转染过pacAd5-Luc-FGF21-3′UTR报告基因的正常小鼠的外泌体转入
AdicerKO
小鼠中,观察到小鼠肝脏和循环血中的
Fgf21 mRNA
都明显减少。之后作者又分别将含有和不含有
miR-99b
的
AdicerKO
小鼠的外泌体转入AdicerKO小鼠和正常小鼠体内,发现转入含有miR-99b的外泌体小鼠其Fgf21 mRNA明显减少。(FIG 4)
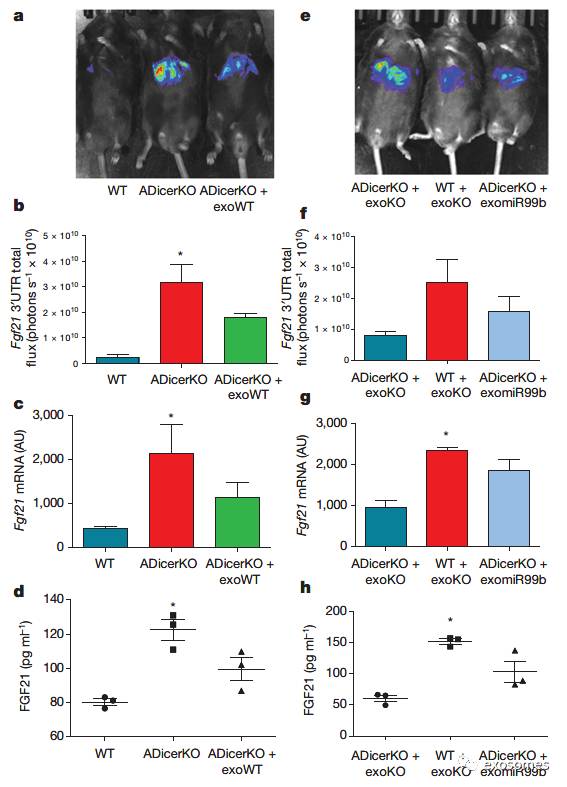
图四:在体内实验中,外泌体miR-99b能调节FGF21。