植物免疫学领域关于PTI
(PAMP-triggered immunity)
和ETI
(Effector-triggered immunity)
的研究是非常重要的问题。
位于细胞膜表面的PRRs可以识别来自于病原菌的PAMPs, 进而诱导宿主植物天然免疫抑制病菌的进一步侵染
【1,2】
;
而病原菌可以通过分泌蛋白因子绕过或者抑制植物的PTI反应;相应地,植物进化出特异性识别效应蛋白的免疫因子,比较代表性的如NLR类受体蛋白,通过直接或者间接的与效应蛋白相互作用,诱发ETI免疫,这一过程通常伴随剧烈的生理活动变化,例如过敏反应
(HR)
【3,4】
。
ETI 介导的免疫通常是针对特异性的病菌,甚至特定的效应因子;
而PTI一般表现为广谱缓和的抗性机制。
但是
ETI反应的基因型特异性,对不同病菌的免疫广谱性尚缺乏系统性的研究。
近日,加拿大多伦多大学细胞与系统生物学系的
Darrell Desveaux
和
David S. Guttman
团队合作在
Science
杂志上发表一篇题为
The pan-genome effector-triggered immunity landscape of a host-pathogen interaction
的研究论文。
该研究利用泛基因组学对丁香假单胞菌与拟南芥互作过程中三型效应因子介导的ETI现象进行了系统研究,发现了新的ETI信号通路,并且对于ETI反应在效应因子识别物种特异性中的重要作用有了新的认识。
该研究首先运用BLASTP对数据库中分布于28个国家来自于100种寄主植物的494种丁香假单胞菌株的基因组序列进行比对分析,分析总结了丁香假单胞菌的三型效应因子的分布情况,并对发现的所有效应因子序列进行分类,经过一系列序列比对优化归为70个蛋白家族, 622个多频次序列基因。
基于三个原则:
1、该效应因子起始密码子上游10kbp内包含保守的Hrp-box启动子序列;
2、该效应因子的序列和Hrp-box上游区域不含有简并碱基;
3、该效应因子的Hrp-box及其上游25bp范围不包含简并碱基,并且不在重复序列的末尾,选取其中529基因中有代表性的效应因子进行基因合成,分别在受体菌株中表达,便于后续ETI反应的表型研究。
进一步对建立的数据库PsyTEC
(
Pseudomonas syringae
Type III Effector Compendium
)
进行评估,研究人员认为本研究中的分析方法已经可以高效的体现丁香假单胞效应因子家族的广泛分布多样性。
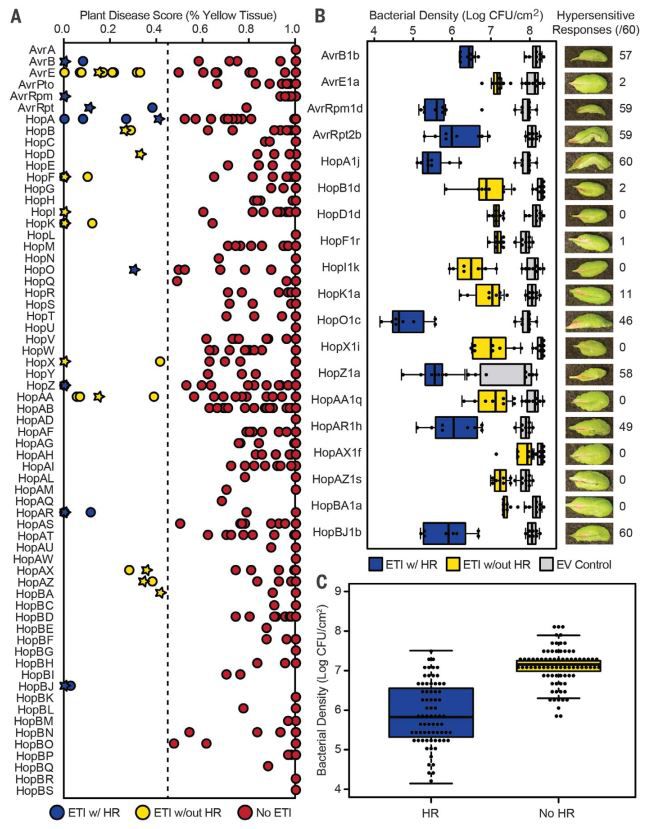
该研究的第二个目的是鉴定到丁香假单胞中所有可以引起拟南芥Col-0 ETI反应的效应因子。将529个PsyTEC
效应因子在DC3000中表达,发现有些基因可能没有正常表达,然后对Col-0进行侵染筛选,筛选结果鉴定到59个可以诱导ETI反应的效应因子,来源于19个蛋白家族,同时不同效应因子的类似表现体现了免疫激发子在不同遗产背景下致病表型的保守性。选取19个蛋白家族中各一个代表检测其在植物上的病菌生长情况,发现含有ETI激发子的病菌生长速率明显下降,这与ETI反应的结果一致。 研究人员同时对鉴定到的ETI激发子关键核苷酸进行突变研究进一步证明了这些蛋白功能完整性对诱导ETI的重要性。HR是ETI反应的典型表型,但是该研究发现有些不明显的HR依然可以作为ETI反应的表型,HR的强度与病菌生长速度下降存在强烈的正相关性。研究人员对理论上广泛存在于全球494种丁香假单胞菌引起的ETI反应进行评估,其中影响评估结果的因素主要有1、不同效应子在不同菌株中表达会影响ETI反应;2、质粒上表达的效应子可能与染色体表达的效应子在诱导ETI功能上有差异;3、每个PsyTEC分枝的效应子仍然存在约5%的氨基酸序列差异,这个差异也可能导致ETI效应的改变。
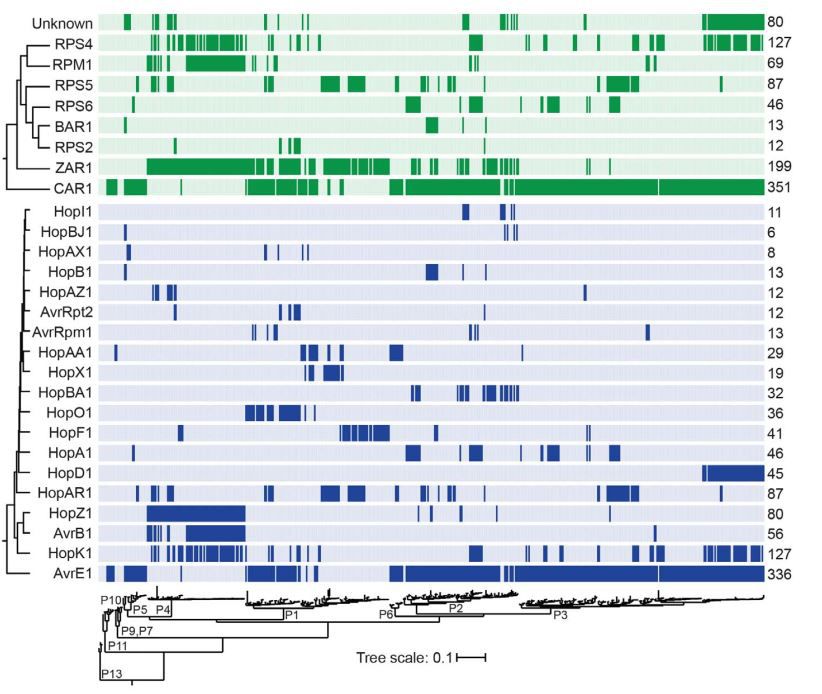
研究人员进一步拟南芥中与19个ETI激发子家族相关的NLRs进行筛选,在DC3000中表达每个ETI激发子然后侵染不同NLR突变体植株,通过表型实验确认ETI反应中的对应关系,最终鉴定出能被所有ETI激发子诱导ETI反应的NLR蛋白。
筛选结果确认了已经报道的所有
NLR—效应子
组合,同时鉴定到两个新的NLR基因:
At5g18360
(Toll/interleukin-1类受体可以识别HopB)
命名为BAR1
(HopB-Activated Resistance 1)
和At1g50180
(coiled-coil NLR类可以识别AvrE和HopAA1)
命名为CAR1
(CEL-Activated Resistance 1)
,通过基因编辑CAR1进一步验证了其识别AvrE和HopAA1的功能。
作者认为可以根据NLRs与ETI激发子之间的对应关系预测并定位NLR基因,该研究还发现,拟南芥几乎可以对丁香假单胞完全免疫,并且仅仅通过极少数的抗性NLRs蛋白介导,八个NLRs抗性蛋白可能识别96.6%的丁香假单胞菌, 同时68%的菌株可以被多个NLRs抗性蛋白识别。
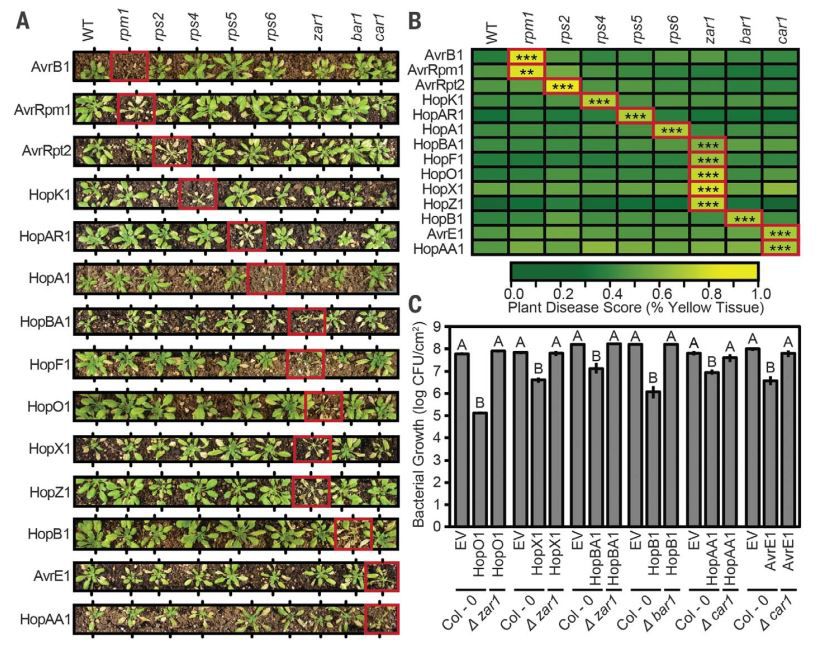
研究人员最后研究了是否单个NLR蛋白介导的ETI反应就可以决定病菌识别寄主的特异性。
对比强毒力菌株PmaES4326和PmaYM7930发现,两者ETI激发子情况的差异主要是是否含有HopAR1,病菌侵染实验中,含有HopAR1的PmaYM7930在Col-0上仅仅表现微不足道的病症且与PmaES4326在植物叶片上病菌生长情况存在两个数量级的差异,而在
rps5
突变体上两种病菌的生长水平几乎一致,致病能力也类似;










