看见五颜六色花里胡哨的图,就走不动了。看这篇有什么新鲜点没,和大家一起成长bigger than bigger~(回复
170523
可下载文献,一月有效)
在肺肿瘤发生过程中染色体12qF1的miRNA表达增加
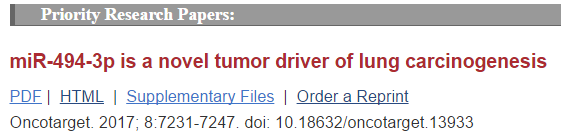
小鼠肺肿瘤代表图像,正常非诱导(N)、它莫西芬诱导的非肿瘤性(4-OHT+N)、肺增生(Hyp)、肺腺瘤(Ad)、腺癌(AdCa)(Fig.1A);
分析显示在肺癌发展过程中大部分miRNAs表达异常(补充图1A);
与肿瘤前、癌性病变组织相比,非肿瘤组织表现出明显不同的miRNA表达谱(Fig.1B);
鼠染色体12qF1簇中大多数miRNA(30/54,55%)在肺组织癌转化过程中过表达(Fig.1C,黑点无改变,红点改变量最大);
通过qPCR证实了染色体12qF1中的miRNA上调(补充图1B)。
已知12qF1中miRNA的表达与小鼠老化相关,接下来按着不同时间段、年龄段从模型小鼠身上取非肿瘤肺组织。数据显示,12qF1的miRNAs不受肺衰老影响,因为,26个miRNAs(48%)在所有样本中均未表达,15个miRNA(28%)在所有标本中都表现出相当水平。在老年组织中,分别仅有9个(17%)和4个(7%)的miRNA,分别减少或者过表达(补充图1C)。
因此,来自K-Ras(+/LSLG12Vgeo);RERTn(ert/ert)小鼠模型肺组织中染色体12qF1簇 中miRNA的上调与肺癌发生相关,排除衰老影响。

14q32 miR-494-3p与肺癌患者预后相关
鼠染色体12qF1 miRNA区域,相应的人类基因组是保守序列,在染色体14q32的DLK1-DIO3位点。
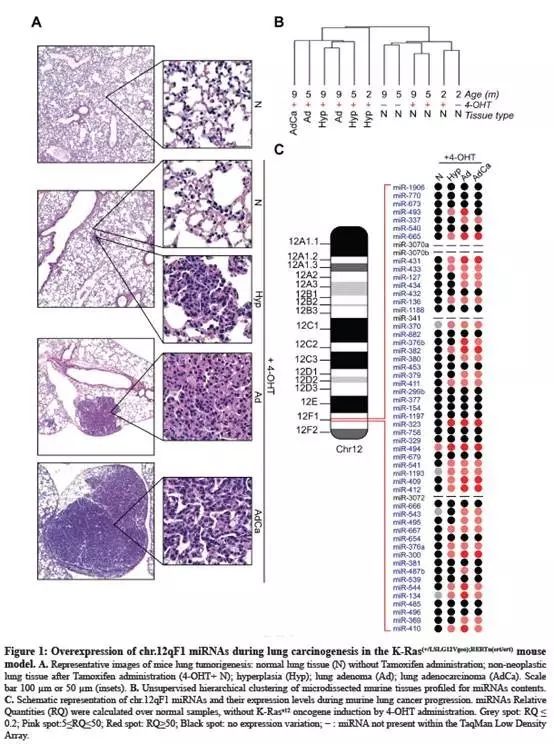
从57例NSCLC患者中,分析一个亚型(n=10)中的miRNA(miR-127、-300、370、-379、-382、-409-3p、-412、-431、-494-3p和-543)。尽管与正常组织相比,这些miRNA在肺癌中一般不会显著表达(附图2A),但是miR-494-3p表达水平较高的患者,其无病生存时间较短(p=0.046;补充图2B);
NSCLC患者扩大样本到113例,miR-494-3p高水平表达患者与较短的总生存期显著相关(p=0.03;Fig.2A);其他miRNA不存在单独与NSCLC患者存活率相关(一个没影响)。然而,14q32染色体中大多数(9/10)miRNA在肿瘤组织中表达的升高与患者无病生存率的缩短存在相关性(多了就影响了,Fig.2B);
研究14q32 miRNAs表达与肿瘤干细胞标志物之间的潜在关系,检查53个肿瘤样本中CD44、ABCG2、ALDH1A1、NANOG和c-MYC基因的表达(Fig.2C和补充图2C)。当肿瘤细胞中14q32 miRNAs共同(>8)上调时,ABCG2的表达量显著上调(Fig.2C);
miR-494-3p在侵袭性前体中的表达明显高于肿瘤体(Fig.2E);
其他miRNA的表达在两个肿瘤区域之间是无法区分的(补充图2D)。
从小鼠肺癌模型和患者样本得出:miR-494-3p和14号染色体miRNA簇与NSCLC中ABCG2-阳性表型具有相关性。

调控miR-494-3p的表达
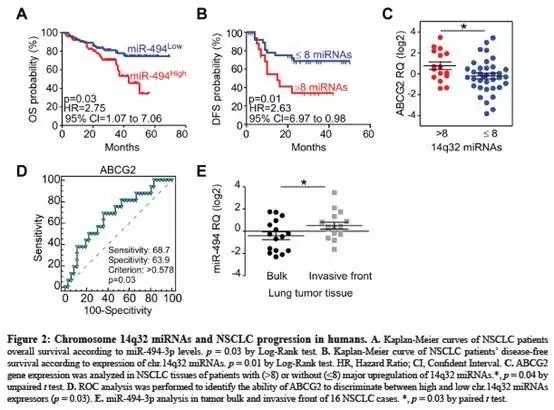
在NSCLC中检测到前体pri-miR-494与其成熟miR-494-3p的表达之间存在直接相关性(Fig.3A);
具有pri-miR-494高表达的肿瘤显示出较高的miR-494-3p水平(Fig.3B);
接下来检测表观遗传学,16CpG位点区域中的基因间甲基化差异(IG-DMR)。正常组织(60.9%)显示比相对应肿瘤(40%)更高的IG-DMR CpG甲基化水平(Fig.3C-D);
相对于非肿瘤性肺组织,肿瘤中IG-DMR去甲基化更为明显,肿瘤细胞中pri-miR-494表达水平明显升高(Fig.3E);
检测了14号染色体33区域的亚显微基因组改变,但该区域不包括DLK1-DIO3基因座(见补充材料),14q基因组与pri-miR-494表达之间没有直接的相关性(Fig.3F);
结果显示:后期的修饰改变可能有助于miR-494-3p在肺癌中的调控,而不是基因层面的改变进行的调控,接下来研究NSCLC中潜在的miR-494-3p相关信号通路。
miR-494-3p调节参与癌症发展过程的通路
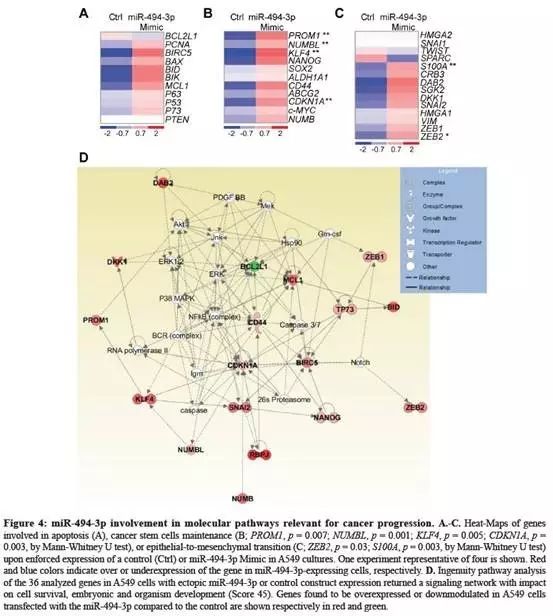
在A549细胞中,分析了受miR-494-3p表达调控的36个涉及重要癌症途径的基因(补充材料);
与对照组相比,miR-494-3p的强制表达不影响细胞凋亡相关机制(Fig.4A);
但是与对照相比,显著增加了PROM1,CDKN1A,NUMBL和KLF4的水平,4个与干细胞相关的mRNA(Fig.4B);
miR-494-3p过表达之后,间质转化因子(EMT)发生上调,ZEB2和S100A的mRNA显著增加(Fig.4C);
通路分析表明:miR-494-3p调节影响涉及“细胞死亡和存活、胚胎发育、生物发育”等细胞调控网络(Fig.4D,红色上调、绿色下调);
miR-494-3p参与肺肿瘤起始种群维持
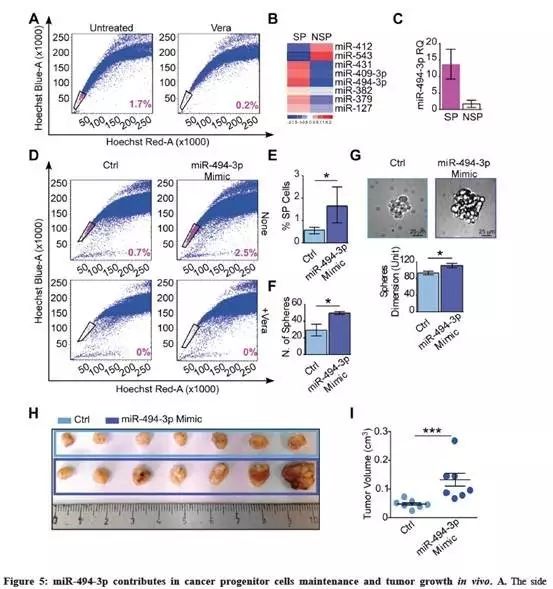
为了更好地了解14q32 miRNAs,特别是miR-494-3p在肿瘤干细胞维持中作用,从A549肺癌细胞分选SP和NSP细胞群体(Fig.5A);
A549细胞中,10个14q32 miRNA中有8个的表达在基线条件下(miR-127、379、382、40-4-3p、-412、-431、-494-3p和-543)(补充图3B);SP细胞与NSP细胞相比表现出明显不同的miRNAs表达谱(Fig.5B);
miR-494-3p miRNA表达差异最高的miRNA(Fig.5C);
在A549中过表达miR-494-3p,并通过流式细胞仪检查SP细胞区域的百分比。与对照相比,较高水平的miR-494-3p导致SP的显著增加(Fig.5D-E);
使用miR-494-3p抑制剂转染A549,降低其表达后,与对照相比并不影响SP比例(补充图3C-D);
与对照相比,非贴壁培养条件下,转染miR-494-3p模拟物表达显著增强了A549球形成能力,增加球型大小(Fig.5F-G);
异种移植检测miR-494-3p在体内维持肿瘤生长中的作用,将转染过表达miR-494-3p/对照miRNA的A549细胞皮下注射到无胸腺小鼠。注射18天后,与对照相比,miR-494-3p过表达组产生较大的肿瘤(Fig.5H-I)。
miR-494-3p过表达增强肿瘤活性
分析miR-494-3p在肿瘤细胞运动和转移中的作用。
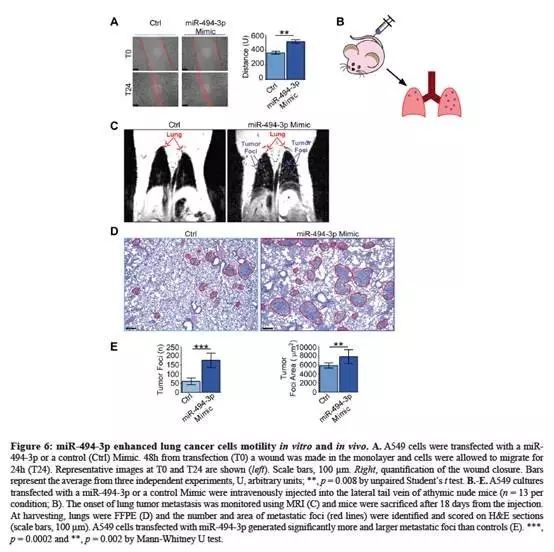
在伤口愈合实验:miR-494-3p过表达后显著增强了A549细胞迁移能力(Fig.6A);在补充实验中还进行Boyden chamber实验。
将A549细胞注射到无胸腺小鼠的外侧尾静脉,在第11天和第17天通过MRI(核磁)监测到肺癌灶形成(Fig.6B-C);
第18天收小鼠肺样本,组织
切片进行评分。与对照组相比,miR-494-3p过表达注射组,小鼠表现出转移病灶数目和表面积显著增加(Fig.6D-E);未检测到肝转移。
miR-494-3p有助于NOTCH1途径和PTEN调节
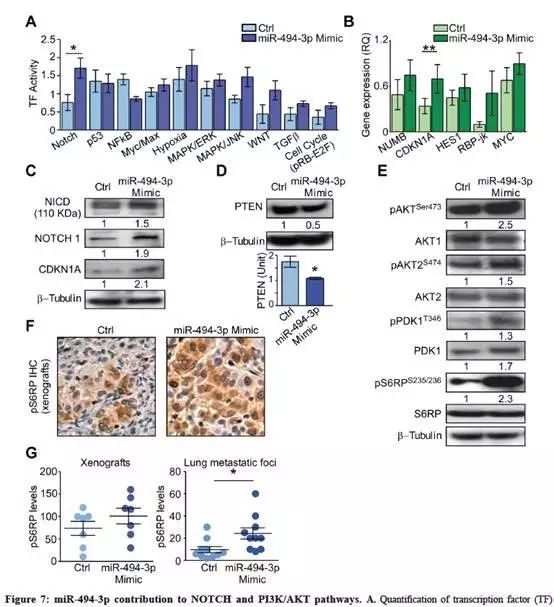
分析10个致癌转录因子,A549-miR-494-3p过表达细胞表现出NOTCH1通路活性显著增加(Fig.7A);
已知的NOTCH1靶基因,CDKN1A和RBP-jk的mRNA也显著上调(Fig.7B);
在蛋白水平,miR-494-3p过表达增加了总NOTCH1、细胞内NOTCH1(NICD)、CDKN1A的蛋白表达量(Fig.7C);NOTCH1激活、PTEN被抑制。
miR-494-3p过表达与PI3K途径活化相关:AKT在Ser473磷酸化,AKT2在Ser474磷酸化、总AKT、AKT激活的PDK1、pS6RP蛋白均显著上调(Fig.7E);
相比之下,miR-494-3p的水平降低并不影响NOTCH1激活、NOTCH1靶标CDKN1A的水平以及A549细胞中PI3K信号传导的水平(补充材料);
体外切片观察到类似结果:miR-494-3p过表达异种移样本,病灶切片显示磷酸化-S6RP染色增加(Fig.7F-G);
实验完了,理一下故事线
miR-494-3p促进PI3K信号传导的激活,同时在肺癌细胞中激活AKT-mTOR-pS6RP,进而助于肿瘤细胞增殖;同时,miR-494-3p可激活NOTCH1信号转导途径以及下游基因(参与EMT和cancer stemness)的表达。
这些结果指示出,一新的miR-494-3p-NOTCH1-PI3K信号转导方式调控肺癌发生和转移的激活。
我收获了两点:1、Fig.4D的做法值得去摸索摸索。2、知道了新概念,miRNA簇。补充图&材料感兴趣的小主儿可以从后台下载,就不再展开了。






