近日,德国马克思普朗克作物育种研究所
Charles J. Underwood
教授研究团队
在国际学术期刊
Nature Genetics
在线发表了题为
“
Harnessing clonal gametes in hybrid crops to engineer polyploid genomes
”
的研究论文(图
1)
。
该研究创建了二倍体番茄不同遗传背景中有丝分裂替代减数分裂
Mitosis instead of meiosis
(
MiMe
)
体系,并通过杂交不同
MiMe
突变体,成功创制了拥有四套无重组的、完整祖父母基因组的四倍体番茄植物。该研究为多倍体渐进式杂种优势
(AHP)
的固定提供了重要理论依据和实践指导价值。该研究首次在植物中实现了来自不同亲本的两个克隆配子的结合。

图1,在番茄中操纵克隆配子体来实现多倍体基因组设计
杂种优势(hybrid vigor or heterosis)是自然界中广泛存在的遗传学现象。它指的是两个不同亲本杂交所产生的子一代在生物量(如产量)、生长率、繁殖力和适应性等方面表现出优于双亲的特征。1776年,德国学者科鲁特(Kolreuter)用烟草属植物杂交,发现其杂交子代具有明显的增产效应。1908年,美国植物与遗传学家沙尔(Shull)首次提出了杂种优势的概念。如今,杂种优势已被广泛应用于粮食和经济作物的育种中,包括玉米、番茄、土豆和水稻等(图2),并且杂交育种取得了令人瞩目的成就。

图2,玉米、番茄、土豆和水稻中的杂种优势。
(Zhang
et al.
, 2021; Springer and Stupar, 2007; Krieger
et al.
, 2010; Zhang
et al.
, 2016)
回顾两百多年的玉米育种历程,可以看到上世纪30年代美国杂交玉米开始商业化,传统的单杂交和双杂交技术推动了杂交种的迅速发展。研究表明,在四倍体玉米中存在多倍体渐进式杂种优势(Allotetraploid Progressive Heterosis, APH) (图3)。然而,由于减数分裂过程中同源染色体上等位基因的重组交换,F1代中的杂种基因型在下一代无法固定,这导致了优良的杂种优势在后代中的分离(图4)。因此,如何有效的固定杂种优势对种业发展起着至关重要的作用。

图3,二倍体和四倍体玉米中的渐进式杂种优势。
(左图来源于https://www.pioneer.com/us/agronomy/domestication-of-corn.html; 右图来源于Washburn and Birchler, 2014)

图4,有性生殖过程中杂种优势在下一代无法固定。(减数分裂过程中同源染色体重组使得亲本遗传物质发生分离
(Wang
et al.
, 2015)
)
2009年6月9日在线发表的文章
(d’Erfurth
et al.
, 2009)
为此项研究奠定了重要的基础,并且在单子叶作物水稻研究中也有了直接应用
(Mieulet
et al.
, 2016)
,但是这些
MiMe
组合(SPO11-1&REC8&OSD1, PAIR1&REC8&OSD1)在双子叶作物蕃茄上却无法直接应用。其中主要原因为OSD1在拟南芥和水稻中存在一个极其类似的拷贝UVI4(主要参与植物营养生长-叶子发育过程中内复制),而在番茄中只有一个拷贝(UVI4突变影响植株下胚轴伸长)
(Di
et al.
, 2022)
。为避开有性生殖中减数分裂重组过程,该研究首先对减数分裂DNA双链断裂形成(前期重组)、减数第一次分裂中姐妹染色单体的共分离以及减数第二次分裂这三个关键事件进行调控,以便创制出
MiMe
三突材料,进而获得二倍体的克隆配子体(固定母本遗传物质)。为了快速获得相关突变体的表型,该研究使用了生长周期短、转化效率高的特殊矮化番茄材料(Micro-Tom)进行转基因验证。通过设计6个串联sgRNA靶位点来敲除参与以上途径的三个重要因子(SPO11-1,REC8和TAM),最后成功获得避开减数分裂重组的
MiMe
突变体。实验结果表明,
Slspo11-1
(消除减数分裂重组)和
Slrec8
(破坏姐妹染色单体之间的粘连)功能缺失突变体都显示雌雄败育的表型,
Sltam
(部分避开第二次减数分裂)单突能产生重组的未减半二倍体雌雄配子体(2n)(图5),并且
Sltam
能够恢复
Slspo11-1
Slrec8
双突中雌雄完全败育的育性,实现将减数分裂转变为类似有丝分裂的可能(图6)。此外,该研究通过高通量花粉测定仪(Mutlisizer 4e),定量分析了
MiMe
突变体中产生二倍体花粉的比例(36.75%-62.18%)。
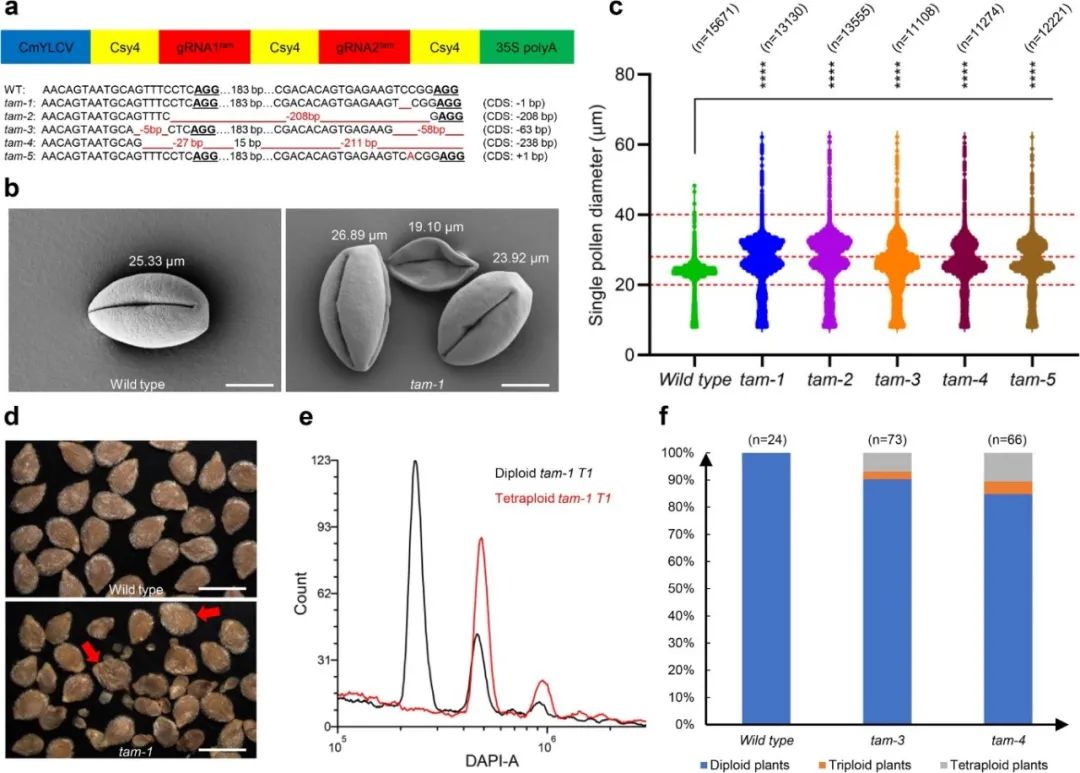
图5,
Sltam
突变体能产生未减半二倍体配子。
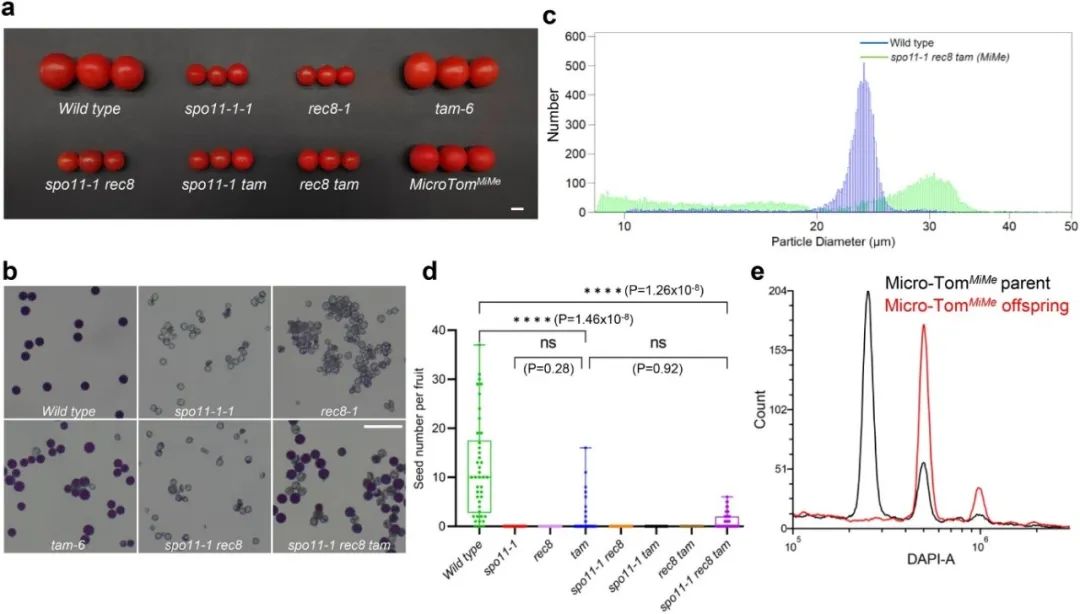 图6,番茄自交系中的
MiMe
系统。
图6,番茄自交系中的
MiMe
系统。
为验证
MiMe
突变体能产生克隆的、无重组的雌雄配子体,该研究将这种
MiMe
策略运用到杂交番茄中,直接开展了在三个不同杂交种(包括两个商业化番茄品种: 德国先正达公司的Funtelle和荷兰Enza Zaden公司的Maxeza)中编辑这三个基因的转基因实验,获得了不同遗传背景的
MiMe
突变体材料。为了证实
MiMe
突变体能产生二倍体克隆配子体,该研究对杂交F1代、F2后代和
MiMe
突变体自交后代进行了表型与基因型分析。全基因组测序结果表明,相比遗传变异大的F2后代群体,
MiMe
突变体产生的四倍体植株没有发生重组, 具有比较一致的表型特征(图7)。更加有趣的是,通过杂交不同的
MiMe
突变体,该研究获得了拥有四套祖父母基因组的全新四倍体材料(图8),该材料能够产生形状正常的无子果实,拥有更多抗病性位点与基因。
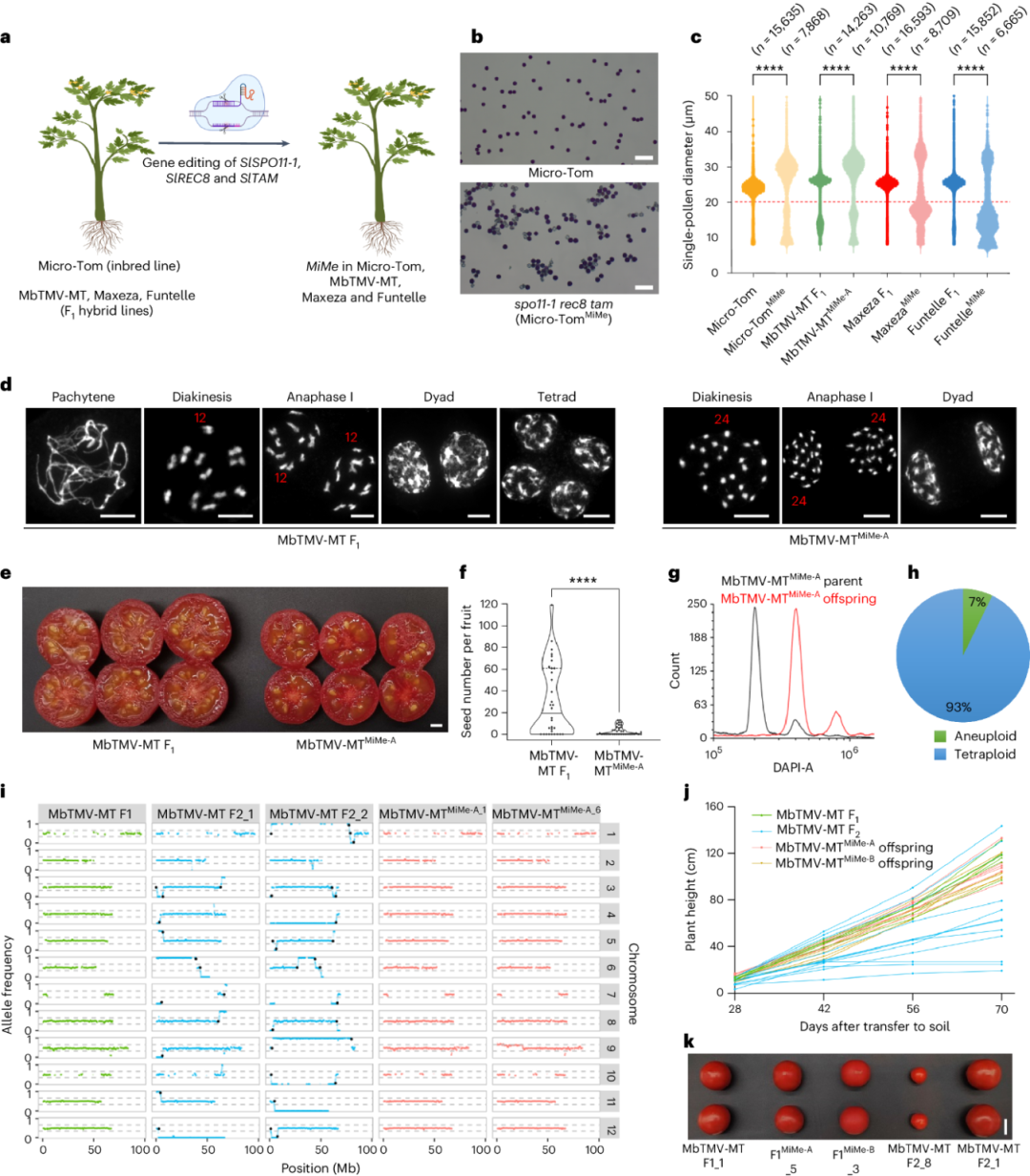
图7,
MiMe
突变体能产生与母本遗传物质一致的克隆配子体。
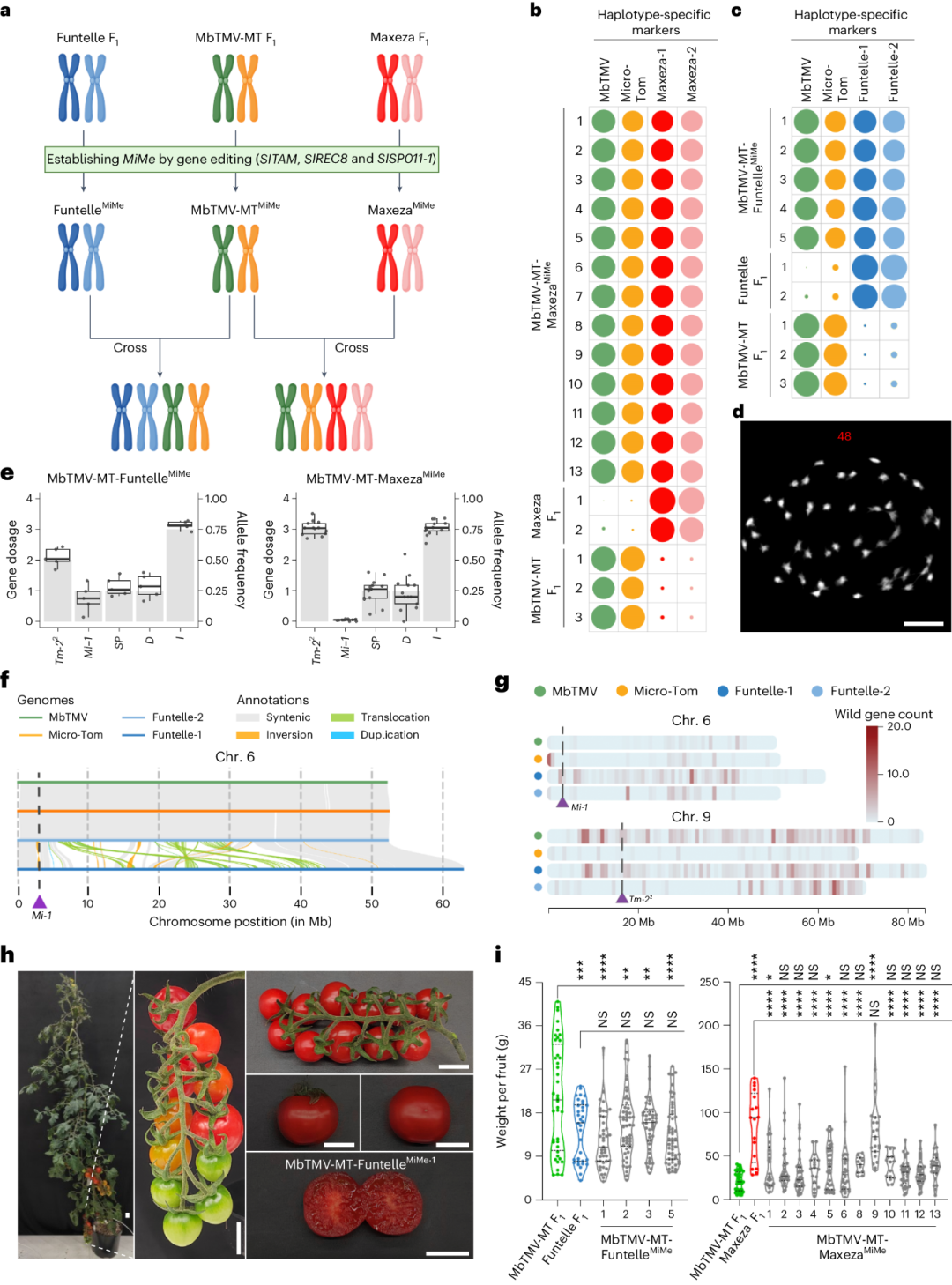
图8,精准设计四倍体番茄植株。
多倍体基因组设计在当前和新型多倍体作物的育种中具有直接的应用潜力。首先,多倍体基因组设计可以有效地将任何杂交育种计划的遗传图谱长度加倍(由二倍体转变为四倍体)。在番茄中,这将有助于通过将野生作物近缘种(与现代栽培种存在200多万年的进化历程)的完整基因组渗入栽培系中来引入非生物和生物抗性。其次,多倍体基因组设计可用于以受控的方式(无重组)利用多倍体植物额外的杂种优势,提供一种在二倍体水平上进行遗传改良的便捷途径,然后在实际育种中利用多倍体水平上的杂种优势,这可能尤其适用于马铃薯(二倍体与四倍体)育种。多倍体基因组设计可能预示着杂交多倍体作物育种新时代的开始,以应对气候变化、疾病压力和人口增长的直接挑战。
德国马克思普朗克作物育种研究所博士后
汪亚中
为该论文第一作者
,
Charles J. Underwood
教授
(
目前任职于荷兰拉德堡德大学
)
为通讯作者。此外,
R
oven Rommel Fuentes
博士后、
Willem M. J. van Rengs
博士
(目前任职于
荷兰
Keygene公司)
、
Sieglinde Effgen
技术
研究员、
Mohd Waznul Adly Mohd Zaidan
博士
(
目前为马来西亚农业研究与发展研究所
MARDI
研究员
)
、
Rainer Franzen
研究员、
Tamara Susanto
在读
博士生、
Joiselle Blanche Fernandes
博士后、
Raphael Mercier教授
为该研究做出了重要贡献。该研究得到了马克思普朗克学会、德国研究基金会
DFG
、欧洲研究理事会
ERC
、德国亚历山大洪堡奖学金、马来西亚农业局奖学金等多个项目的资助。此外,根据相关研究成果,Charles J. Underwood
教授、汪亚中博士后和
Raphael Mercier
教授已申请欧专利
No. 23179909.9
一项。
该项研究发表后,2024年5月21日Nature Genetics
上线了由汪亚中博士后和
Charles J. Underwood
教授受邀撰写的
题为“克隆配子使多倍体基因组设计成为可能”
的研究简报
:

专家评论
:
“作者通过将四种不同的单倍型依次组合成四倍体植物,描述了一种番茄同源多倍体渐进杂种优势的系统。
该系统的重要性在于,与传统育种相比,它可以实现更大范围的等位基因组合和更多的杂种优势。
”
——美国加利福尼亚大学戴维斯分校Venkatesan Sundaresan(著名植物繁殖生物学家、 美国国家科学院院士)
编委评价
:
“
这项研究提出了一种在杂交番茄基因型中生成克隆配子的方法,通过四种预定义基
因组单倍型的受控组合,实现了多倍体基因组设计。令人激动不已的是,当我们看到
有丝分裂替代减数分裂系统被引入番茄,这种建立的渐进式杂种优势框架对植物育种
可能具有潜在的应用价值。”
——《自然遗传学》高级编辑Wei Li
参考文献
:
d’Erfurth, I., Jolivet, S., Froger, N., Catrice, O., Novatchkova, M. and Mercier, R.
(2009) Turning Meiosis into Mitosis G. P. Copenhaver, ed.
PLoS Biol.
,
7
, e1000124.
Di, S., Zhang, P., Zhang, J., Liu, G., Wang, G., Shi, Q., Ma, F. and Bao, Z.
(2022) Tomato UVI4 homologue modulates cell expansion to participate heat-stimulated hypocotyl elongation.










