
从感情上不愿意相信随机突变能产生复杂的结构,是攻讦进化论最常见的原因。比如许多人看到眼睛的解剖结构都会不由自主地问:什么样的突变能从从零开始,把顽肉改造成这样精密的光学系统?
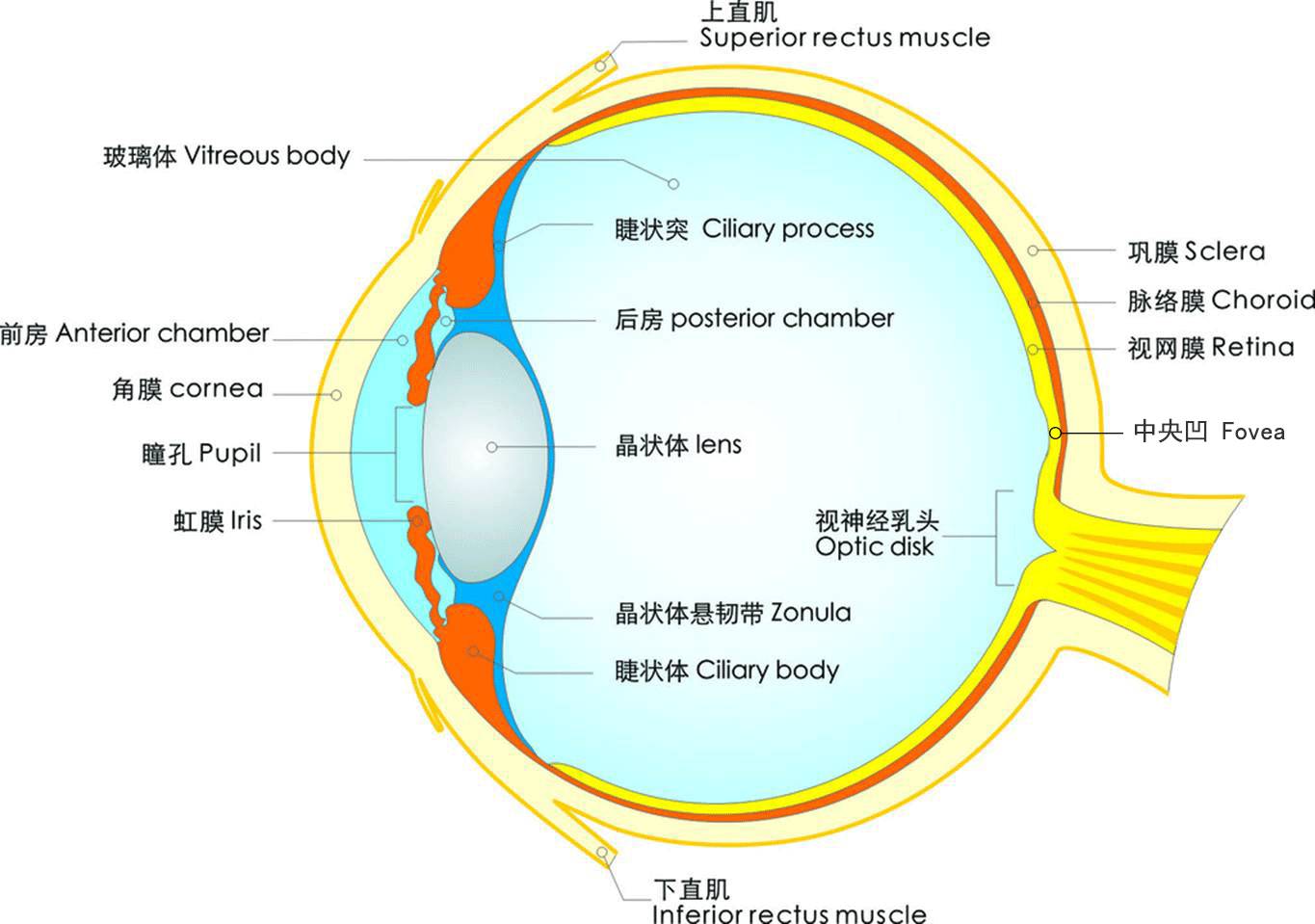
人眼球的解剖
这种困惑首先来自一种误解,即认为感受光刺激是一种“特异功能”,感光细胞和普通细胞有巨大的差异——其实不然,任何生物需要感受光刺激,只需富集色素即可,不同的色素会吸收不同频率的光,将其转变为化学能。
比如人眼的视杆细胞就用视紫红质感光,甚至可以在黑暗处捕捉到单个光子;而视锥细胞用三种不同的色素构建了三原色的彩色视觉。
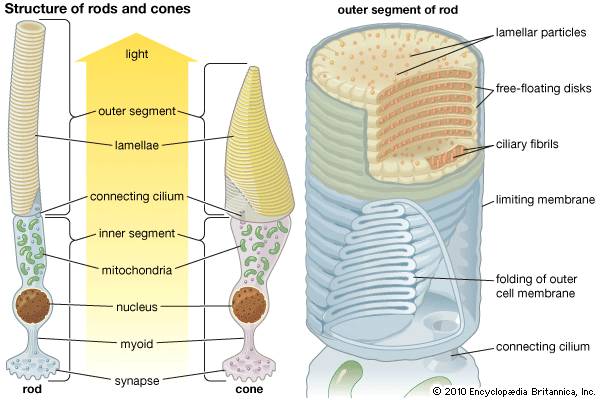
人眼的感光细胞,相比普通细胞,它们最显著的特征就是有厚厚一大摞膜结构,上面满是感光用的色素
另一个最常提到的例子是眼虫。这类单细胞生物属于古虫界,既不是动物也不是植物,既能光合作用也能主动摄食,它们的鞭毛基部粘有一小团类胡萝卜素——自然界最常见的色素——受鞭毛遮挡,这团色素会因为不同的朝向吸收不同剂量的光能,反馈为鞭毛的摆动方向,最终帮助眼虫游向光亮的地方,促进光合作用。这个功能非常类似眼睛,所以那团类胡萝卜素就被称为“眼点”。
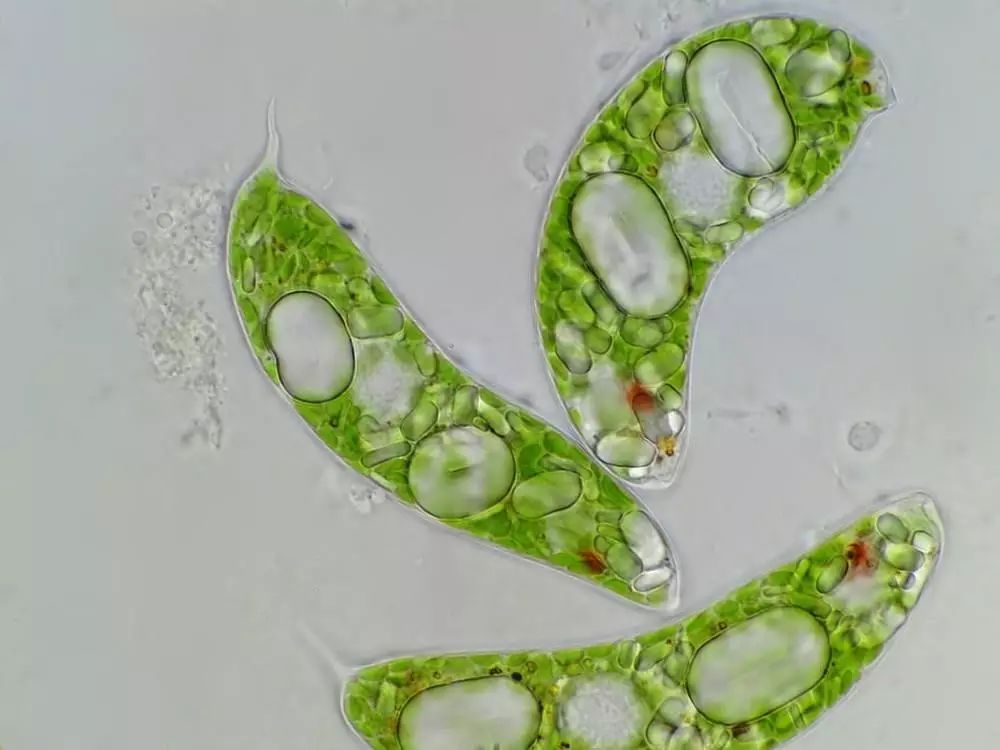
三只眼虫,绿色的胶囊状小颗粒是叶绿体,白色的卵状大颗粒是光合作用积累的淀粉,红色斑点就是眼点
其次的一个误解,是认为“人进化出了人的眼睛”,然后质疑人眼周围是毫不透明的肌肉和骨骼,不可能突变出光通路需要的透明组织,但眼睛的进化历史要远远长于人类这个物种的进化历史,甚至长于脊椎动物的进化历史——在遗传学和发育学的研究中,我们找到了一些关键的基因,比如 PAX6 基因。
这个基因源自所有两侧对称动物的共同祖先,在神经系统和眼睛的发育中发挥着关键作用;而且高度保守,哺乳动物的 PAX6 基因可以在昆虫身上发挥一样的功能,这意味着动物界 30 多个门的眼睛在进化的极早期有相同的来源:一个覆盖色素的凹陷,并没有成像功能。

扁形动物门的三角涡虫,它的眼睛非常接近我们共同祖先的眼睛:就是两个铺有色素的凹陷,一些神经细胞伸入凹陷,感受光刺激引发的化学反应

左边是我们看到的图像,右边是三角涡虫看到的图像
但这是一个好的开始,凹陷不但能保护脆弱的色素,更能大致区分光线的来源,这对趋利避害很有帮助。在此基础上,动物界用 5 亿多年的时间,平行发展了 40 到 60 个谱系,进化出了许许多多更加精致复杂的眼睛,尤其是脊索动物门的脊椎亚门、软体动物门的章鱼目和节肢动物门的昆虫纲,代表了视觉进化的三个顶峰——当然,也共用着高度相同的 PAX6 基因。
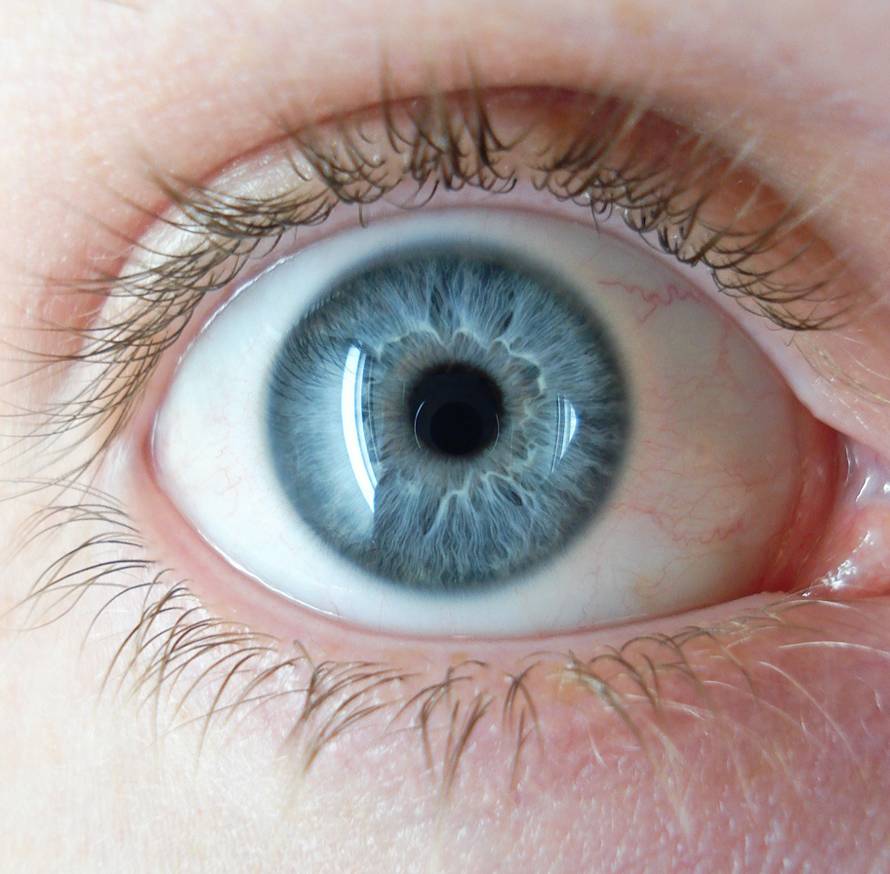
脊椎动物亚门,人的折射单眼

章鱼目,北太平洋巨型章鱼的折射单眼

昆虫纲,雄性四节蜉蝣的三只反射单眼(淡蓝色)和四只折射复眼(橙色和褐色)
问题集中于脊索动物门,我们眼睛的早期形态在文昌鱼身上体现得最多,也研究得最充分——文昌鱼虽然被称为“鱼”,但它属于头索亚门,远比鱼所在的脊椎亚门更接近脊索动物门的祖先,是难得的活化石物种。
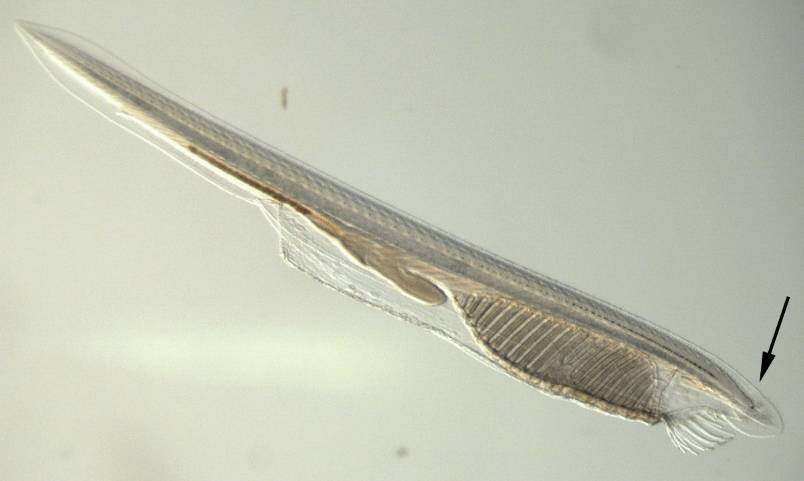
文昌鱼,箭头所指的小黑点是“额眼”
文昌鱼身体含水量很高,非常透明,有一条卷入体内的神经索贯穿头尾,这条神经索两侧分布着许多感光细胞,其中含有视黑蛋白,对蓝光敏感。而脊椎动物的中枢神经系统,包括大脑和脊索,在胚胎发育上也都源自这条卷入体内的神经索,只是发达得多。视黑蛋白在我们身上与生物钟的昼夜节律有关。
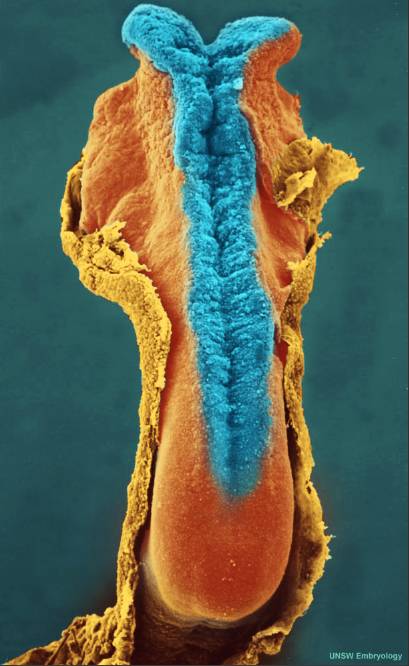
妊娠四周的人胚胎,背部的细胞(神经板)正在向内卷(蓝色),它最终将会发展为中枢神经系统
特别的,在文昌鱼的神经索前端背面,受 PAX6 基因控制,还有一个较大的带色素的杯状凹陷,里面分布了两列感光细胞,我们称为“额眼”(Frontal eye)。这个结构与我们的双眼在进化上同源,其中的感光细胞与我们的视锥细胞和视杆细胞同源,带色素的凹陷与我们的视网膜上皮同源。
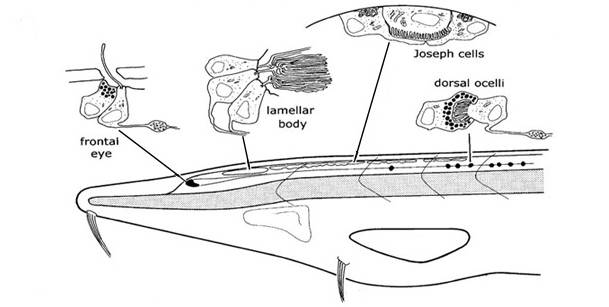
文昌鱼的几组感光器官,包括额眼(Frontal)、板层小体(lamellar body,可能与我们的松果腺同源)、约瑟夫细胞(Joseph Cell)和背单眼(dorsal ocelli)
也就是说,脊索动物的眼睛在进化早期随着神经索的发育卷入了体内,不再暴露于体表,但当时的组织高度透明,原始的眼睛在体内仍可以感受光刺激。此外还要特别注意,这个内卷还反转了眼睛,使得左边的感光器官要穿透组织看右侧,右边的感光器官要穿透组织看左侧。脊椎动物的早期胚胎中也是一样,即将发育为眼睛的凹陷来自内卷的神经管,左边朝右,右边朝左。
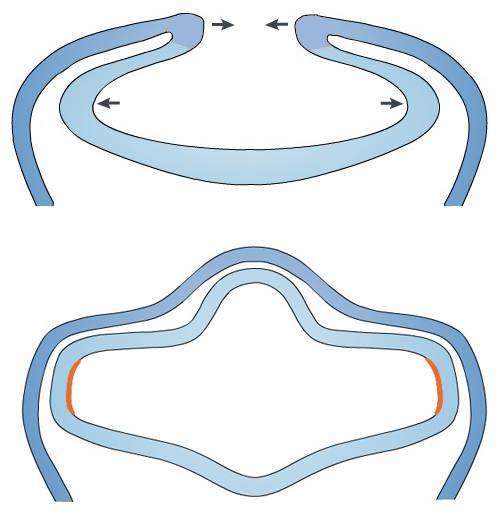
神经板内卷成神经索,外层蓝色代表体壁;内层浅蓝色管道代表卷入体内的神经管,将来发育成中枢神经系统;橙色代表神经管两侧将要发育成眼睛的凹陷
通过对脊椎动物胚胎发育的进一步研究,比如对斑马鱼胚胎发育的研究,我们发现,随着组织越来越不透明,脊椎动物再也不能左眼看右,右眼看左,这对将要发育成眼睛的凹陷还要再次翻转,而且随着翻转程度加深,一部分体壁上的细胞会填入凹陷,发育为角膜、玻璃体、晶状体等屈光结构。
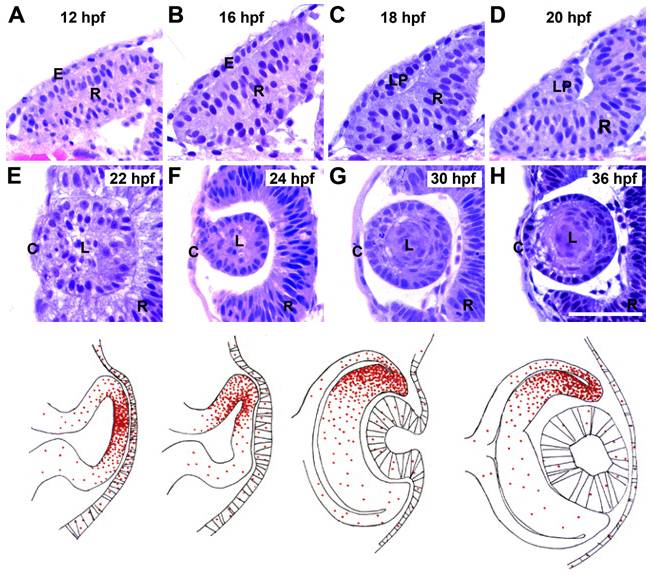
斑马鱼胚胎中眼睛的发育历程,上方为显微照片,下方为示意图,可见原本朝向左侧的凹陷逐渐翻过来朝向右侧,卷成碗状,同时开口处的细胞移入碗中,发展为晶状体等屈光结构




