正文
近
日,
GPB
在线发表了中国医学科学院
“
本草基因组协同创新团队
”
首席专家宋经元课题组题为
“Comparative Genome Analysis of
Scutellaria baicalensis
and
Scutellaria barbata
Reveals the Evolution of Active Flavonoid Biosynthesis”
的研究论文。我们的
“
要文译荐
”
栏目很高兴邀请到共同第一作者高冉冉博士为大家解读《中华人民共和国药典》收载的唇形科黄芩属药用植物黄芩和半枝莲的基因组及其利用研究。
黄芩属药用植物全基因组解析及其黄酮类成分生物合成进化机制研究
联合二代及三代测序技术、
Hi-C
染色体构象捕获技术等破译黄芩属药用植物黄芩与半枝莲染色体水平基因组;通过
OrthoFinder
方法对候选物种的单拷贝直系同源基因进行鉴定并构建系统发育树揭示物种起源及进化;基于同源基因的同义替换速率研究揭示候选物种的分化及全基因组复制事件;通过
PFAM
、
BLASTP
等方法注释及挖掘黄酮类成分生物合成相关基因;基因组共线性及系统进化分析揭示候选基因的进化机制。
发现黄芩和半枝莲的分化时间远早于之前的报道,鉴定唇形科与胡麻科共享的全基因组复制事件。
揭示了黄芩和半枝莲黄酮合成途径的趋异进化,如PAL和CHS在黄芩中特异的串联复制,4CL在半枝莲中特异的扩张。
鉴定了黄芩与半枝莲物种形成后黄酮羟化酶(F6H和F8H)的串联复制及功能分化情况。
黄芩(
Scutellaria
baicalensis
)和半枝莲(
Scutellaria barbata
)均为唇形科黄芩属药用植物,黄芩的干燥根及半枝莲的干燥全草是常用中药,具有清热解毒的功效。黄芩属约有
300
–
360
个物种,唯有黄芩和半枝莲被《中华人民共和国药典》收载。新型冠状病毒肺炎(COVID-19)正肆虐全球,中医药在新冠诊治方面发挥积极作用,清热解毒药——黄芩受到关注。国家中医药管理局发布的《新型冠状病毒肺炎诊疗方案(试行第七版)》,由黄芩等21味中药材组成的“清肺排毒汤”可用于轻型、普通型及重型新冠肺炎患者的治疗。基于《新冠诊疗方案(试行第七版)》《中医疫病名方精选》推荐治疗新型冠状病毒肺炎恢复期的中医方剂,通过分子对接VEGFR 和FGFR 靶点,筛选出潜在抗肺纤维化活性成分命中率30%以上的中药,其中以黄芩最多。黄芩和半枝莲的主要活性化合物为黄酮类成分,包括黄芩素、野黄芩素、汉黄芩素及其糖苷类化合物。最新药理研究显示黄芩提取物具有有效的抗新冠病毒(SAR-CoV-2)活性,黄芩素及其类似物是强效的SARS-CoV-2 3CLpro抑制剂。

黄芩

中药是我国的战略资源,药用植物是中药的主要来源
,
由于中药基原植物遗传背景
不清、优良品种匮乏等因素,导致中药材质量良莠不齐,很大程度上影响中医用药安全
有效。因此,利用组学技术揭秘中药基原植物基因组遗传信息具有非常重要的意义。本研究联合采用二、三代测序及
Hi-C
技术,绘制黄芩和半枝莲高质量基因组图谱,挖掘黄酮类成分生物合成途径相关基因,通过比较基因组研究揭示黄芩和半枝莲黄酮类成分生物合成及进化机制,为中药黄芩和半枝莲活性成分合成生物学及优良种质选育等研究奠定基础。
黄芩基因组组装大小为
377.0 Mb
,
contigN50
为
2.1 Mb
,
99.8%
的序列被挂载到
9
条染色体上(
2n = 18
),共注释出
33,414
个蛋白编码基因和
2833
个非编码
RNA
(
ncRNA
);半枝莲基因组组装大小为
353.0 Mb
,
contigN50
为
2.5 Mb
,
98.8%
的序列被挂载到
13
条染色体上(
2n = 26
),共注释出
41,697
个蛋白编码基因和
1768
个
ncRNA
。两个物种的基因组组装质量均较高,可以满足目前基因挖掘、物种进化、基因组重测序等下游分析。黄芩与半枝莲之间基于基因的共线性分析揭示其染色体数目变异和结构重排(
图
1
)。此外,在
DNA
序列水平上的比对也显示了黄芩和半枝莲基因组存在大规模的结构变异。
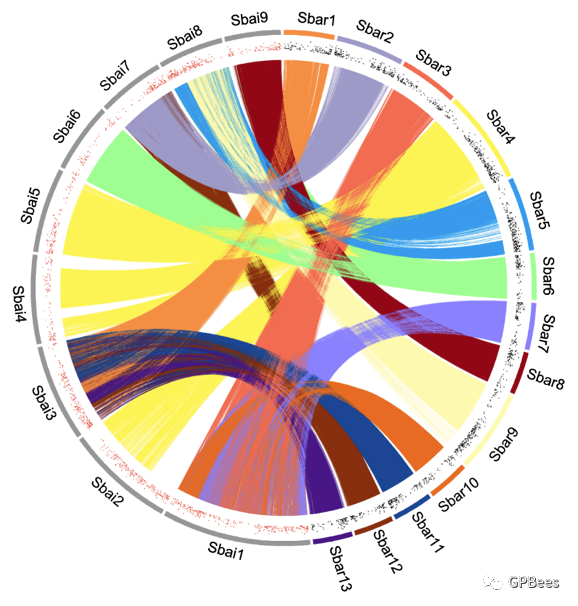
图
1
黄芩和半枝莲基因组共线性分析
黄芩和半枝莲基因组分别鉴定出
1225
和
1654
个长末端重复序列(
long terminal repeats, LTRs
)插入,且
LTRs
的插入时间黄芩要早于半枝莲(
141
万年前和
88
万年前),推测近期
LTRs
的插入是导致半枝莲染色体扩张和重排的关键因素。此外,黄芩和半枝莲基因组分别注释出
142,951
和
147,705
个简单重复序列(
simple sequence repeats, SSRs
),可用于后期黄芩和半枝莲的遗传多样性及分子育种研究。
通过单拷贝基因串联构建系统发育树揭示唇形科黄芩属药用植物与同科鼠尾草属药用植物丹参亲缘关系最近,其分化时间为
41.01
百万年前(
million years ago
,
MYA
);黄芩与半枝莲分化时间为
13.28 MYA
,远早于之前报道的
3.35 MYA
;唇形科与胡麻科分化时间约为
49.90 MYA
。序列同源性分析显示黄芩与半枝莲之间有
17,265
对直系同源基因,同义替换速率(
K
S
)为
0.16
,代表了黄芩与半枝莲的分化时间;黄芩、半枝莲、丹参和芝麻基因组分别鉴定出
7812
、
7168
、
6984
、
7711
个旁系同源基因,
K
S
值约为
0.87
、
0.86
、
1.02
、
0.67
。进化分析显示全基因组复制(
Whole genome duplication
,
WGD
)事件发生在这
4
个物种分化之前,唇形科和胡麻科物种共享的
WGD
事件大约发生在
46.24
–
60.71 MYA
,黄芩、半枝莲与丹参分化之后没有再发生
WGD
事件。黄芩和半枝莲基因组分别与祖先双子叶植物核型(
ancestral eudicot karyotype, AEK
)和葡萄基因组的共线性分析进一步证实了黄芩、半枝莲与芝麻物种分化之前的
WGD
事件(
图
2
)。
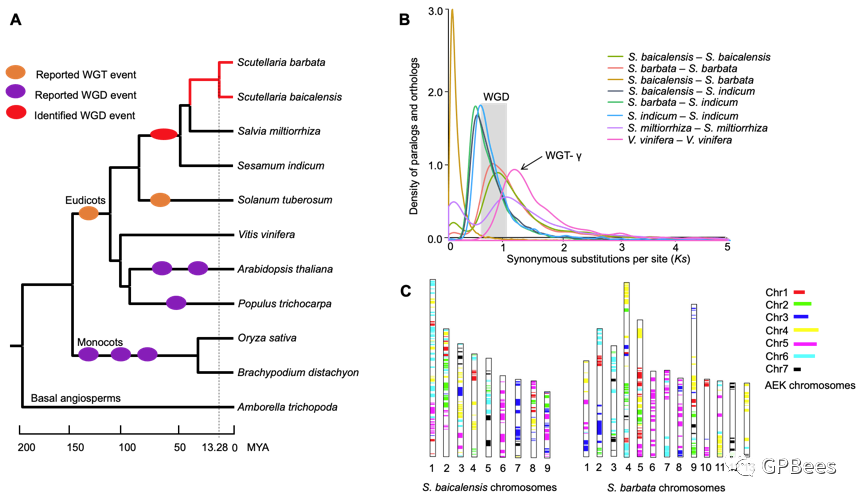
图
2
黄芩和半枝莲基因组的全基因组复制事件分析
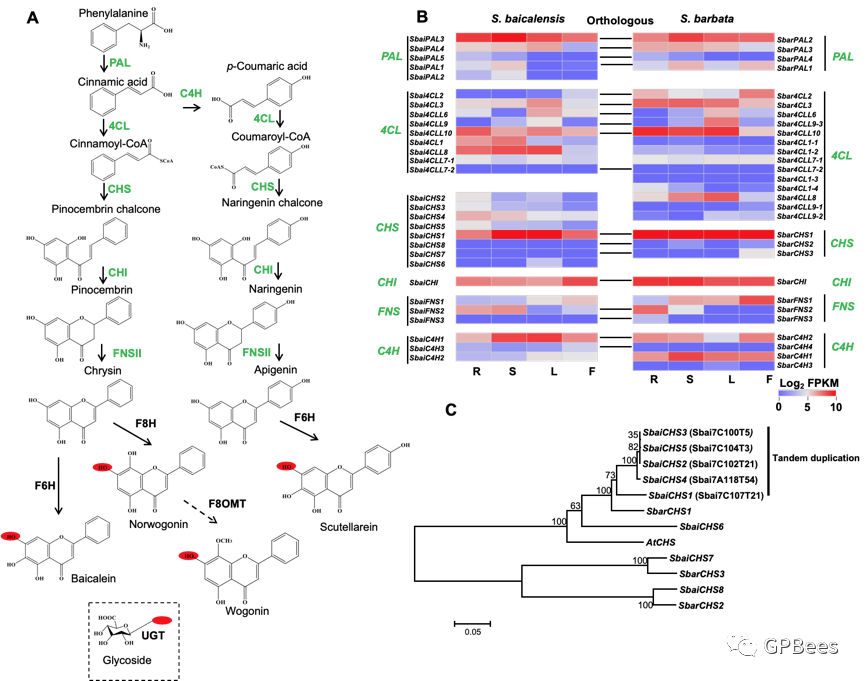
黄芩和半枝莲主要活性化合物为黄酮类成分,包括黄芩素(苷)、去甲汉黄芩素(苷)、汉黄芩素(苷)、野黄芩素(苷)等,利用
UPLC
分析黄芩与半枝莲不同组织部位的黄酮类成分,发现黄芩素(苷)、去甲汉黄芩素(苷)、汉黄芩素(苷)主要在根中积累;野黄芩素(苷)主要分布在茎、叶和花中(
图
3
)。根据黄芩和半枝莲基因组及转录组数据,我们对黄酮类化合物生物合成途径关键酶编码基因进行系统筛选(
图
4
):苯丙氨酸解氨酶(
PAL
)(
5/4
)、
4-
香豆酰辅酶
A
连接酶(
4CL
)(
9/14
)、肉桂酸
-4-
羟化酶(
C4H
)(
3/4
)、查尔酮合成酶(
CHS
)(
8/3
)、查尔酮异构酶(
CHI
)(
1/1
)和黄酮合酶
II
(
FNSII
)(
3/3
),其中
PAL
和
CHS
在黄芩中扩张,
4CL
在半枝莲中扩张,提示在两物种分化后不同基因通过串联复制发生了一定程度的扩张。野黄芩素合成途径关键酶
C4H
在黄芩属地上部位高表达,与野黄芩素的合成与积累呈正相关。
4’-
脱氧黄酮类化合物(黄芩素、汉黄芩素及其糖苷类化合物)在黄芩属不同组织中的表达模式与其化合物的分布也一致。
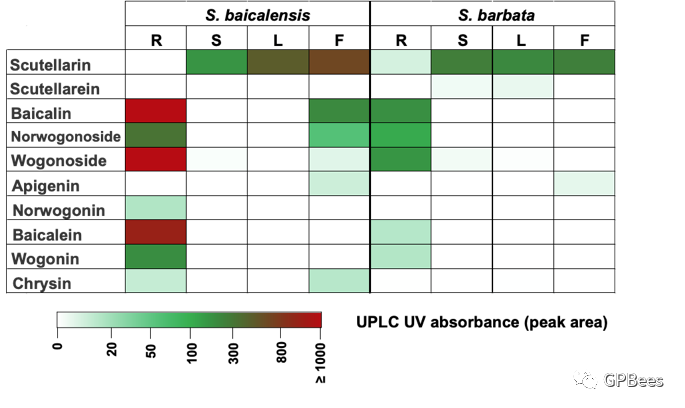
图
3
黄芩和半枝莲不同组织部分黄酮类化合物的特征性分布
黄酮羟化酶
F6H
和
F8H
催化底物白杨素和芹菜素产生黄芩属物种主要的活性成分黄芩素、野黄芩素等化合物。黄芩和半枝莲中分别鉴定出
418
和
398
个
CYP450
基因,各有
9
个
CYP82D
基因(
SbaiCYP82D1-9
和
SbarCYP82D1-9
),可能参与黄芩与半枝莲中白杨素和芹菜素的羟基化反应。已报道
SbaiCYP82D1
具有
F6H
功能,以白杨素或芹菜素为底物生成黄芩素和野黄芩素;
SbaiCYP82D2
具有
F8H
功能,催化白杨素生成去甲汉黄芩素。结合基因表达与进化树分析,推测半枝莲中
SbarCYP82D5
和
SbarCYP82D9
可能具有
F8H
功能,
SbarCYP82D6
和
SbarCYP82D8
可能具有
F6H
功能。基因组共线性分析发现黄芩和半枝莲物种特异的
F6H
功能相关的
CYP82D
基因的串联复制(
SbaiCYP82D1-SbaiCYP82D7-SbaiCYP82D8
,
SbarCYP82D1-SbarCYP82D6-SbarCYP82D8
),以及黄芩和半枝莲物种分化后的
F8H
功能相关的串联复制(
图
5
)。
CYP82D
基因在黄芩属物种中由串联复制导致的基因扩张可能引起黄酮合成相关羟化酶的功能分化。