自噬是一个吞噬自身细胞质蛋白或细胞器并使其包被进入囊泡,并与溶酶体融合形成自噬溶酶体,降解其所包裹的内容物的过程,大隅良典也靠着自噬获得了诺奖。
所以,今天要给你讲的这篇文章,其实跟自噬没什么特别大的关系,讲的是个自噬调控分子。这篇文章发表在Cellular Signalling上(IF=4.191),上了4分的文献告诉你蛋白互作要怎么做。

首先要给你灌输的一个柯霍氏法则,啥是柯霍氏法则?其实简单点说,去掉了某个因素之后,原有的功能丧失,就可以证明这个因素是实现这个功能的关键。
啥?没懂?
好的,给你讲个故事,你就明白了。从前有个专家,做了个实验,他拿了一只蜘蛛放桌子上,他叫了一声“跳!”,然后蜘蛛跳了一下。接着他把蜘蛛的腿都卸掉了,又叫了一声“跳!”,然后蜘蛛动也没动。于是他记录道:蜘蛛的耳朵长在腿上。没毛病……
好了,看完这个,你对柯霍氏法则是不是有了个概念?
那我们接着看这篇文章哈(
警告,以下部分速读太快,请系好安全带,并点击下面的“发车”,然后观看,会让你有超速、压线、逆行的快感
):

摘要部分,我就截取了这么一段,告诉你他们大概做了撒花,一共是三大块:1)共定位;2)蛋白互作;3)凋亡表型。

那他们用了什么样的实验来证明呢?我帮你们总结了一下,大致来说,做了上面这些实验,主要也是为了摘要中的三大块服务的。

接着我们可以看到共定位部分,两个蛋白要相互作用,肯定要共定位一下。起码这俩蛋白需要在一起出现,才能相识相恋相爱是吧,总不能在文章里说这俩蛋白通过粒子纠缠式的网恋,完全是柏拉图式的爱情。但边上为啥要做突变?呃,很简单,还记得刚才的蜘蛛腿么?A蛋白只能抓住B蛋白的腿,但是把B蛋白的腿卸了,A蛋白就注定孤独一生了。
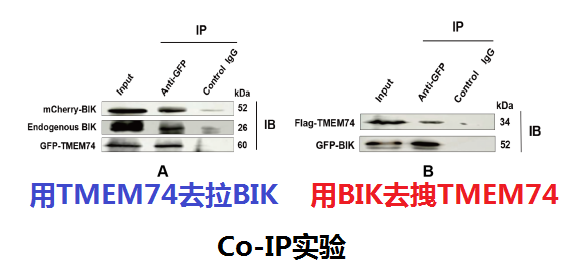
接着他们做的是Co-IP实验,有的人可能不知道啥是Co-IP,那首先我们要简单讲一下IP,这个比较简单,就是先在A蛋白上加上一个标签,然后用抗体去拽这个标签,把A蛋白吸在一个大球上,因为A蛋白能和B蛋白结合啊,那B蛋白势必也会被拉下来,这就是IP。那Co-IP呢?其实就是反过来在用B蛋白拉一遍A蛋白,这两个IP合在一起,就被称为Co-IP。
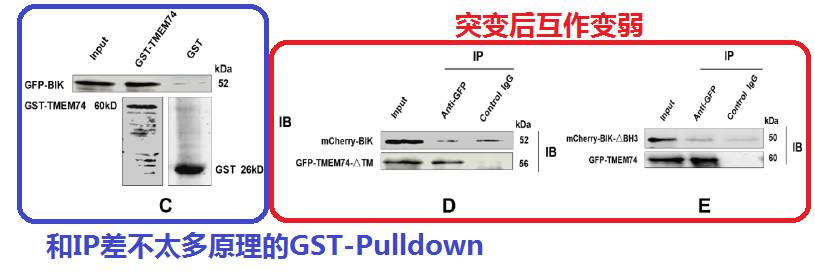
接着是Pulldown,其实原理跟IP差不多,IP是互相拉过表达的蛋白,Pulldown就是拉内源性的细胞自身表达出来的蛋白了。接着还是刚才的蜘蛛腿实验,证明一下突变后,互作变弱了。

说了半天,终于要到表型了,一个是凋亡表型,一个是自噬表型,这里全部先由明星分子代替了。可以看见作为自噬促进剂的TMEM47可以有效抑制凋亡。
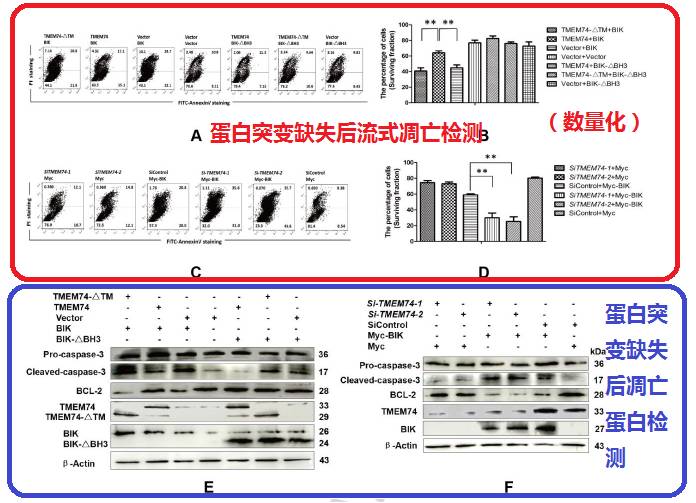
最后用流式再验证了一下凋亡表型,这个时候可以证明的是TMEM47可以抑制凋亡,但这和BIK有啥关系呢?于是把刚才论证过的TMEM47或BIK突变后两者互作变弱,就做了一下这样的验证,分别突变这俩蛋白后,凋亡表型的变化是什么。然后发现,确实变弱了,于是证明TMEM47通过和BIK的互作,抑制细胞凋亡。





