真核生物细胞基因组中的基因需要经过“剪”和“接”这两步化学反应去除内含子,才能令编码区可以连接成不同的信使RNA(mRNA)。近期清华大学的施一公教授在Journal of Molecular Biology杂志上发文,介绍了剪接体近期的重要进展,提供了外显子连接的新线索。
剪接体作为真核细胞中催化pre-mRNA剪接过程的执行者,是真核生物最基本的分子机器之一,对于正常生命活动具有至关重要的作用。但是自上世纪70年代后期RNA剪接的发现以来,科学家们一直在步履维艰地探索其中的分子奥秘,期待早日揭示这个复杂过程的分子机理。
施一公教授研究组通过优化了蛋白提纯方案,首次获得了酿酒酵母U4/U6.U5 tri-snRNP复合物原子分辨率的结构(清华施一公院士发表2016开年《Science》文章),并且捕获了酵母剪接体分别处于激活状态和第一步催化反应后的总体分辨率分别为3.5和3.4埃的两个高分辨率冷冻电镜结构(清华施一公院士Science同日发表两篇文章)。这无疑是这一研究领域的突破性进展。
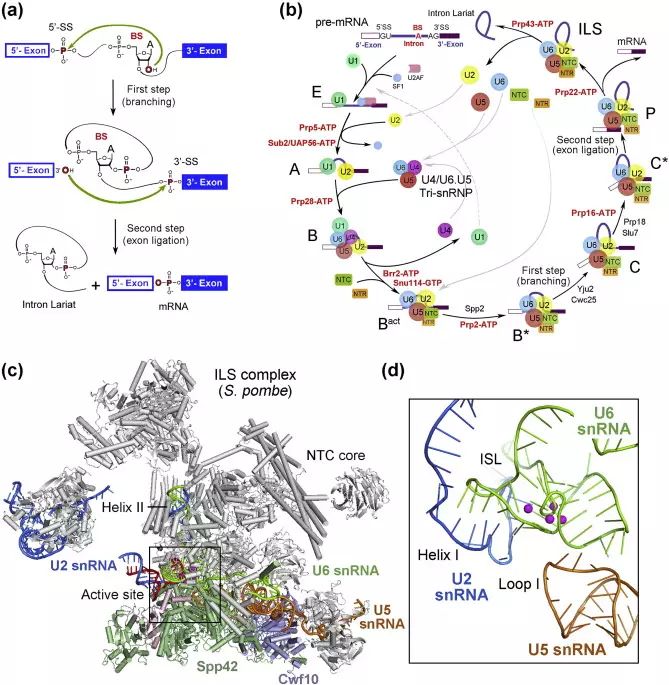
但剪接过程并没有这么简单,剪接体在pre-mRNA剪接反应过程中作为核酶催化需要完成两步转酯反应,第一步是释放5’末端外显子,生成一个内含子-3’末端外显子的套索结构,然后连接上两个外显子,切除内含子。第二步是通过催化激活的剪接体(C*
complex)完成最后进程。
去年年底,这一研究组再次发文,报道了酵母剪接体C* complex,总体平均分辨率高达4.0 Å的冷冻电镜结构。第一步催化反应后的剪接体被称为C
complex,将变化为C* complex的剪接体与第一步剪接体做比较,研究人员发现了一些结构上的变化,如第一步中的剪接因子Cwc25 和 Yju2
已经从活性部位解离了下来,套索结构连接处为即将进入的3’末端外显子序列挪出了15-20
Å的空间等等。
这些结构与之前报道的系列结构组合在一起,组成了一个几近完整的剪接体循环的分子机制拼图,讲述了酵母剪接体的一个完整故事。
然而与酵母剪接体相比,以人类为代表的高等生物的剪接体组成、组装和调控更为复杂,其结构研究也因为组成的复杂性和构象的不稳定性而进展缓慢。由于超过1/3的人类遗传病与剪接过程中的错误直接相关,解析人源剪接体的结构不仅能帮助理解剪接反应的化学本质,更能促进对一些疾病发病机制的理解,并为研发针对剪接体的相关药物提供可能。因此人源剪接体的结构解析是极其重要并亟需解决的难题。
时隔不到半年,施一公研究组又再次公布了人源剪接体的高分辨率三维结构:研究组利用修饰过的pre-mRNA,在体外进行人源剪接体的组装,把剪接反应锁定在了第一步反应之后与第二步反应之前的状态,即C*状态。由于人源剪接体非常不稳定,研究人员使用化学交联剂在温和的条件下对剪接体进行固定,成功获得了稳定的人源剪接体样品,并采用单颗粒冷冻电镜重构出了3.8埃的近原子分辨率结构。
在该结构中,剪接体核心区分辨率高达3.0-3.5埃,清晰地展示了由20余个蛋白与RNA组成的催化反应中心的结构。同时,他们观察到与第二步反应密切相关的剪接因子所呈现出的特定构象,对于稳定反应活性中心以及催化第二步转酯反应至关重要。该结构的解析为揭示第二步反应过程中剪接体的构象变化以及3
'剪接位点的识别提供了重要的结构依据。
原文标题
The Spliceosome: A Protein-Directed Metalloribozyme












