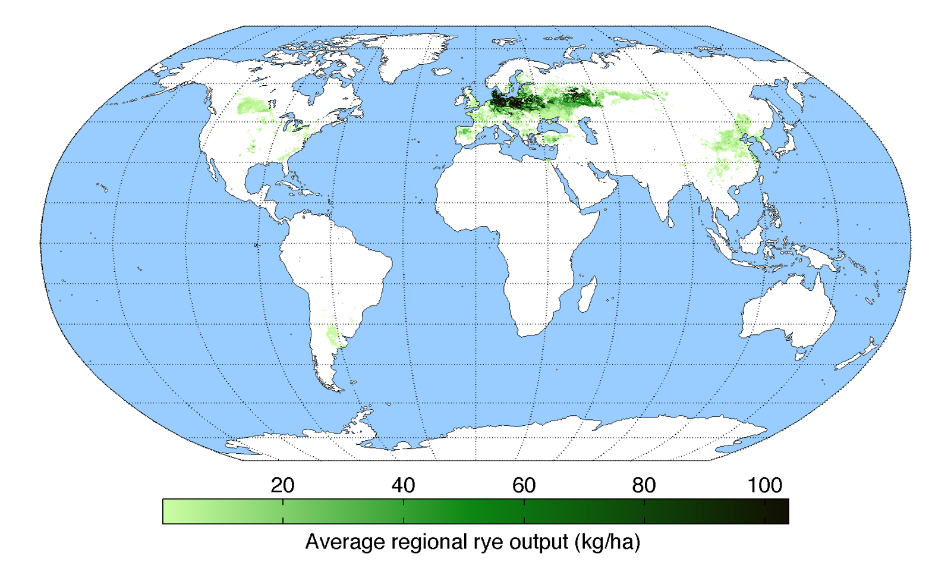
黑麦(
Secale cereale
)是禾本科小麦族黑麦属二倍体植物,是小麦、大麦和燕麦等其他麦类作物的近缘物种。黑麦具有突出的抗寒、抗病、抗逆和耐旱能力,对贫瘠生存环境有很强的适应能力,可耐受其他谷物无法耐受的极端气候条件,对肥料及农药需求较少,种植黑麦可
给自然条件恶劣地区带来显著生态和经济效益。黑麦在欧洲至少已有
7000
年栽培历史,至今依然是北欧地区的主要粮食和饲料作物之一,
黑麦在我国西北、东北和西南山区也有种植
【
1
】
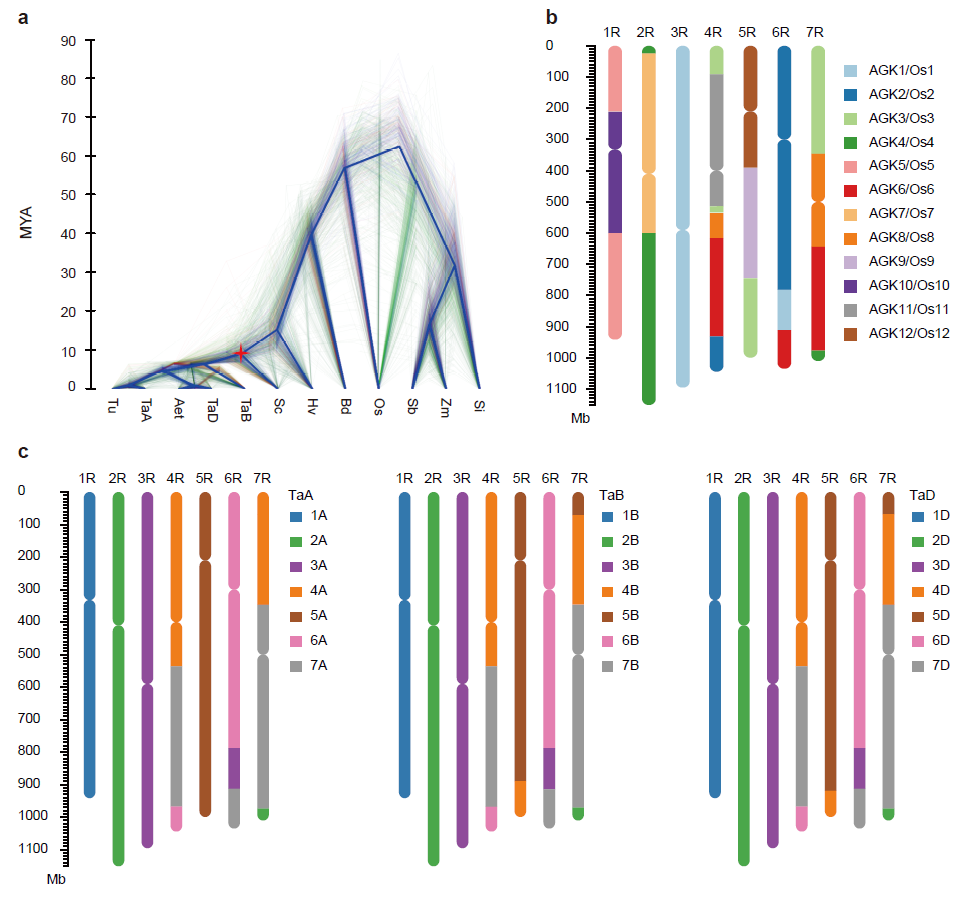
。在全球范围内,黑麦每年有
412
万公顷的种植面积,产量约为
1120
万吨,其中
欧洲占全球黑麦产量的
80
%
(
图
1
)。
图
1
.
全球黑麦产量分布图。
俄罗斯著名
的含酒精饮料格瓦斯和大列巴面包(俄语黑麦面包
хлеб
的音译名字)都是以黑麦为
主要
原料制作而成。写进高中生物教材的
“
小黑麦
”
,是由
二粒小麦
(
AABB
)
或普通
小麦(
AABBDD
)和黑麦(
RR
)远缘杂交
人工合成的异源六倍体或八倍体新物种。通过远缘杂交,黑麦染色体还能部分地替换掉小麦的染色体片段,产生稳定遗传的小麦品系,为小麦育种导入优异外源基因。由黑麦和小麦远缘杂交产生的
1BL/1RS
易位系,能显著提高小麦的抗病性和产量,为保障我国和世界小麦生产以及粮食安全起到了至关重要的作用。据研究,我国栽培的冬小麦品种,
50%
以上都携带有
1BL/1RS
易位染色体,该类品种在北方冬麦区和黄淮冬麦区中占据主导地位【
2
】。
高质量参考基因组序列是生物学功能研究的基础。相对于其他二倍体麦类基因组,黑麦基因组更加复杂(
2n = 2x = 14, RR
)
。前期研究表明黑麦基因组大小为
~
8G
b
,远大于水稻(
25
倍)、谷子(
20
倍),高粱(
11
倍)、玉米(
4
倍)、大麦(
1.5
倍)等其他谷物基因组
、
其基因组中有超过
90%
的序列由高度重复的转座子(
TE
)序列组成。自然情况下黑麦是异交作物
,具有自交不亲和性,其基因组有很高的杂合度。以上这些复杂性是国际上长期以来无法获得黑麦基因组精细物理图谱的主要障碍,
这严重限制了黑麦重要农艺性状遗传基础的解析和禾本科作物比较基因组学研究的深入开展,对黑麦、小黑麦和小麦等重要作物遗传育种改良研究也十分不利
。
为了解决上述科学问题,河南农业大学农学院联合四川农业大学、北京大学和百迈客生物科技公司等多家单位组建了协同创新研究团队,利用中国栽培品种“威宁黑麦”严格自交可结实的特性,得到了高度纯合的材料,运用高通量二代测序、
PacBio
单分子测序、染色质三维构象捕获
(
Hi-C
)
、单分子光学图谱(
Bionano
)和高密度遗传图谱等技术,联合优化多种组装策略,成功构建了威宁黑麦基因组的高质量精细物理图谱(
图
2
)。
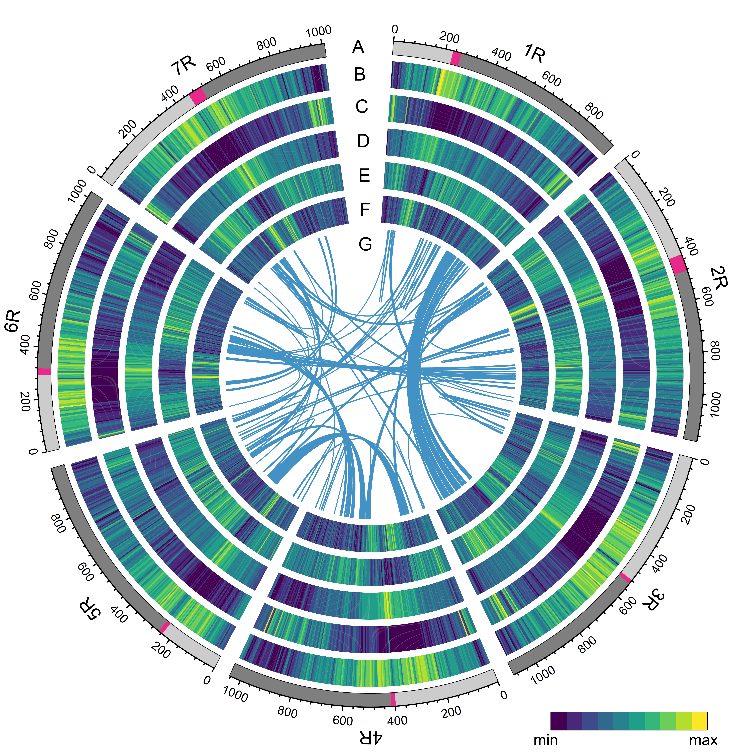
图
2
.
威宁黑麦基因组的重要特征。
(A)
染色体与着丝粒(
红色标注)
,
(B)
转座子
(TE)
密度,
(C)
基因密度,
(D, E)
Gypsy
(D)
和
Copia
(E)
反转录转座子密度,
(F) GC
含量,
(G)
基因组内同源区段。
威宁黑麦基因组组装大小为
7.84Gb
,其中
93.67%
的
序列可以锚定到
7
条染色体上,
有
5
条染色体大小超过了
1Gb
,最大的染色体长度(
2R
)
为
1.15Gb
,
其大小为整个
基因组的
3.6
倍。威宁黑麦中一共注释到
了
45596
个高可信度基因,这些基因中有
1989
个抗病基因。分析发现,黑麦中有
6.99Gb
的
TE
序列
,占基因组比例的
90.31%
,显著高于其他麦类作物。长末端重复反转录转座子(
LTR-RT
)对黑麦基因组扩张贡献最大
,相对于大麦,黑麦多出
2.52Gb
的
LTR-RT
序列,贡献了基因组扩张的
85.42%
,
黑麦中
LTR
有两次明显的活跃复制时间
,一次大约发生在
170
万年前,与大麦
TE
扩张时间相近;更近的一次大约发生在
50
万年前(
图3
)。
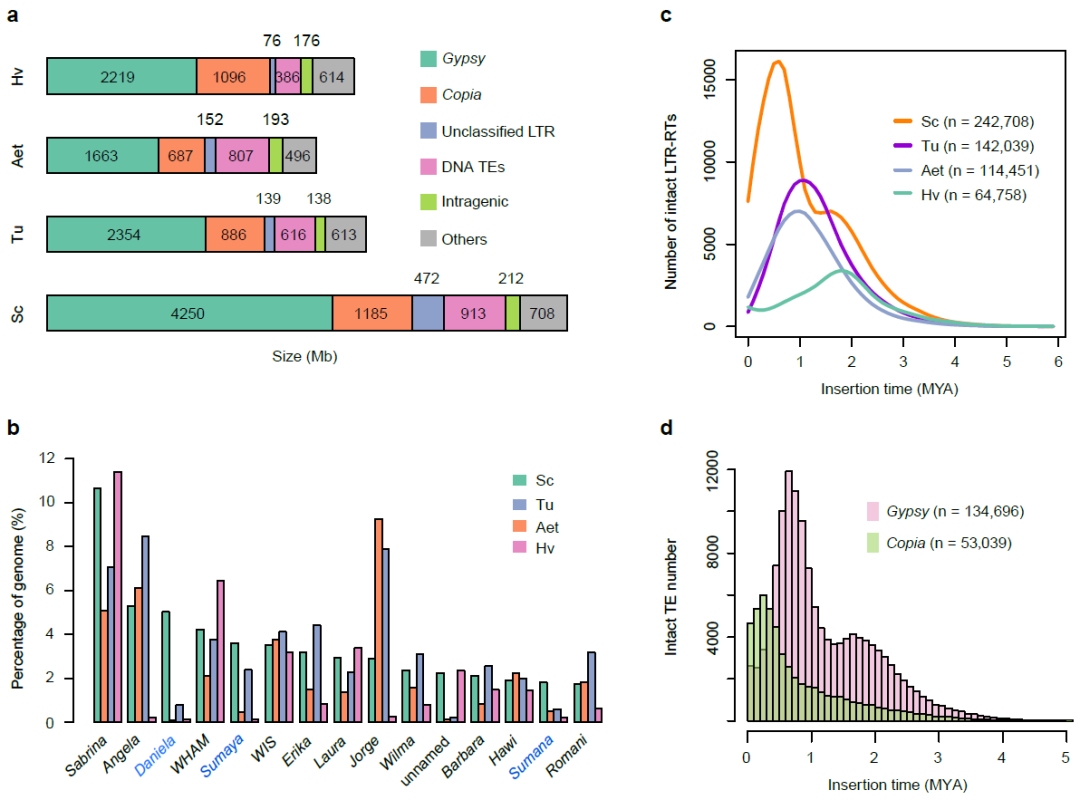
图3.威宁黑麦基因组转座子分析。
研究人员重新
构建了禾本科系统演化史,发现黑麦与小麦共同祖先的分化时间大致在
960
万年前(
图4
a
)。以水稻和小麦为参考,进行比较基因组学分析发现,黑麦基因组结构变异频繁,存在很多特异的染色体重排和大片段易位事件(
图4
b
)。本研究得到的黑麦高质量精细基因组物理图谱,是已知组装完成的最复杂的二倍体禾本科物种基因组。黑麦基因组精细物理图谱的获得,填补了禾本科物种基因组信息的重要空缺,为作物基因组学研究和重要农艺性状功能解析提供了非常珍贵的信息资源。