来源:《中国中药杂志》网络首发时间:
2020-07-28
论中药
“
辨状论质
”
之辨色泽与
品质评价的生物学内涵研究
刘天睿,金艳,孟虎彪,赵玉洋,
周骏辉,袁媛
通信作者
,黄璐琦
(中国中医科学院 中药资源中心 道地药材国家重点实验室培训基地)
中药品质的形成与评价是保障中医药学科和产业发展的关键,但由于中药药效物质的复杂性和功效主治的多样性导致中药品质评价面临巨大挑战。一方面,传统经验鉴别模式依靠眼看、手摸、鼻闻、口尝、水试、火试等方法,依赖专业人员的丰富经验,缺乏客观性指标;另一方面,以指标性成分检测为核心的中药质量现代评控体系,在充分评控中药内在品质上存在一定的局限性,导致经验认知与标准化检验之间存在着鸿沟,中药品质的评价亟需从思维方式、科学理论、方法体系等方面在更高层次的守正创新。
谢宗万、金世元先生先后提出了
“
辩状论质
”“
五象七原
”
等理论,认为药材在
“
形、色、气味、质
”
等外观上所表达的固定特征与其内在品质具有一定的联系,因此可通过对中药材外观性状的分析,作为中药真伪优劣判断的依据。本课题组依据现代生命科学研究成果及发展趋势,提出了道地药材可表现为药材的
“
优形
”
(公认药材性状)和
“
优质
”
(独特化学成分组成),并体现为药材使用上的
“
优效
”
(优于非道地药材的临床功效)。道地药材
“
优形、优质
”
特征的提出,拓宽了传统药材辩状的范畴,其核心思想是通过获取高质量、可重复的性状数据,进而量化分析基因型和环境互作效应及其对中药质量的影响,为中药材现代化质量控制体系建立奠定基础。
色泽是中药
“
辩状
”
的主要内容之一,其包括药材表面的色泽特征、内部颜色以及炮制前后的色泽变化,在中药经验鉴别中具有重要的地位。从生物学上来看,植物颜色的形成是受多种因子共同作用的结果,但其细胞中的色素是颜色形成的根本原因,而研究表明大部分色素具有一定的药理活性。因此,药材的
“
外在色泽优形
”
与
“
内在色素优质
”
是相互统一的。本文拟以
“
辨色论质
”
切入点,系统介绍
“
优色、优质
”
的表征、生物学成因等研究进展,并对色素定量表征在中药质量控制中的应用、
“
优色
”
生物学内涵研究前景进行展望,以期指导建立具有中医药特色的中药质量评价方法。
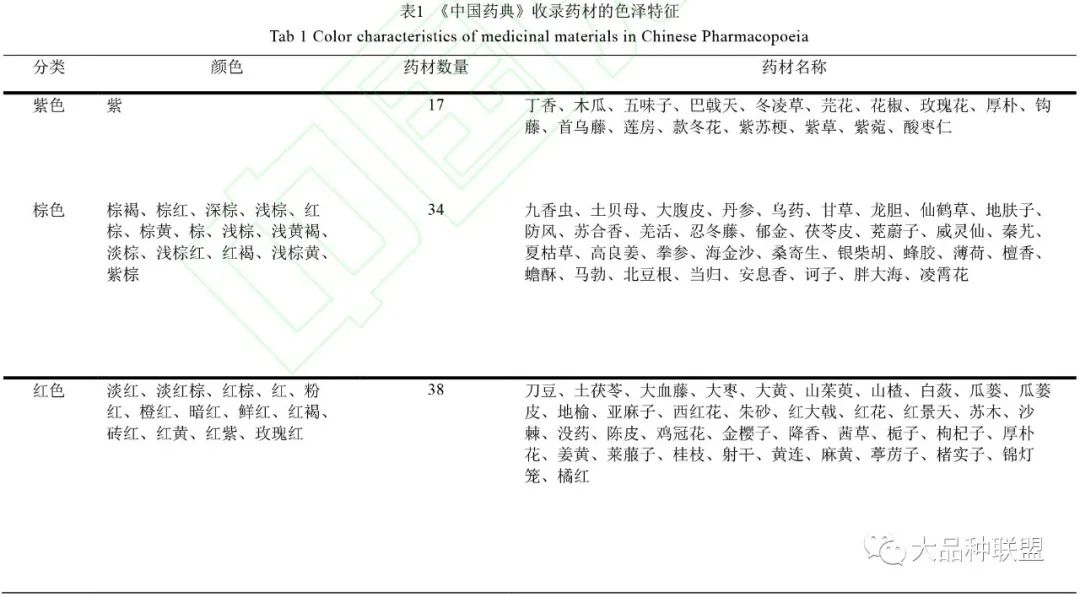
在《中国药典》
2015
年版中,收录了
200
余种药材使用包括色泽等指标进行质量评价,如三颗针外皮灰棕色、大血藤断面皮部红棕色、大枣红色、山茱萸红色等,见表
1
。按照颜色可大致分为紫色、棕色、红色、灰色、黄色、黑色、绿色、白色、其他
9
类,其中以绿色最多,占
18.8%
;其次为黄色,占
18.4%
。第三为红色,占
13.7%
。

在对药材颜色数值进行定量表征研究中,目前多采用
CIELAB
色度空间技术,其规则为:
①
L
表示颜色亮度,即当
L
=0
时,为黑色;当
L
=100
时,为白色;
②
a
表示红绿色轴,即当
a
<0
时,为绿色;当
a
>0
时,为红色;
③
b
表示黄蓝色轴,即当
b
<0
时,为蓝色;当
b
>0
时,为黄色;
④
E
表示所测物颜色的总色差,即
E
=(
ΔL
2
+
Δa
2
+
Δb
2
)
1/2
。随着色度学分析方法的运用,越来越多的研究结果表明,中药的色泽与生物碱、黄酮、萜类等活性成分含量具有一定的相关性。如北豆根药材粉末橙红色越深、总色差越大,且总色差与蝙蝠葛碱含量呈极显著的相关性;色度值可用来反映红花中羟基红花黄色素
A
黄酮类成分的含量;丹参根部越红、活性成分含量越高,其中迷迭香酸、丹酚酸
B
及丹参酮
Ⅰ
与色度
L
值呈极显著负相关,隐丹参酮、丹参酮
ⅡA
与
a
值和
b
值呈现极显著正相关;甘草根皮和断面的颜色均与有效成分含量具有显著或极显著相关性。从上述结果可以看出,颜色
L
,
a
,
b
数值可作为药材质量的评价指标,在此基础上可利用颜色数值表征对药材的品种,采收时期、贮藏、炮制工艺等进行评价,见表
2
。



生物中的色素是一类选择性可将特定波长的光吸收或反射的物质,可见光透过三棱镜可呈现出
7
种颜色组成的光谱,其中红色光波最长,为
640
~
780nm
;紫色光波最短,为
380
~
430nm
,而白色光是一种由二波长光或者三波长光混合而成复合光。中药的色素物质主要包括黄酮类、萜类、生物碱等化合物,在特定条件下可形成红色、蓝色、紫色等,目前大部分中药的色素物质生物合成骨架途径均比较清楚。
2.1
花色素苷
花色素苷又名花青素,是一类糖基化的多酚类化合物,广泛存在于花、种子、果实和营养组织中。常见的花色素苷有天竺葵色素(
pelargonidin
)、芍药色素(
peonidin
)、矢车菊色素(
cyanidin
)、牵牛花色素(
petunidin
)、锦葵色素(
malvidin
)和飞燕草色素(
delphinidin
)等,是多种花色素苷合成酶基因共同表达的结果。花色素苷的生物合成途径己较为清楚,大体分为三个阶段:第一阶段是苯丙氨酸在苯丙氨酸解氨酶
PAL
、肉桂酸
-4-
羟化酶
C4H
和香豆酰辅酶
A
连接酶
4CL
作用下生成香豆酰辅酶
A
;第二阶段是由香豆酰辅酶
A
和丙二酰辅酶
A
经过查尔酮合酶
CHS
催化形成了查尔酮,查尔酮接着在查尔酮异构酶
CHI
的催化下生成柚皮素,然后柚皮素又在黄烷酮
-3-
羟化酶
F3H
、类黄酮
-3’-
羟化酶
F3’H
、类黄酮
-3’5’-
羟化酶
F3’5’H
的作用下生成多种二氢黄酮醇。第三阶段是二氢黄酮醇在二氢黄酮醇还原酶
DFR
的催化下生成了各种无色的花色素,然后在花白素双加氧酶
LDOX
和尿苷二磷酸葡萄糖类黄酮
3-O-
葡萄糖基转移酶
UFGT
作用下将无色花色素转化为有色花色素苷。
花青素合酶(
ANS
)是花色素苷生物合成的关键酶,
SmANS
在白花丹参中过表达可增加其花色苷的积累,同时减少了丹酚酸
B
和迷迭香酸的合成,并恢复了紫红色表型
[54]
。铁皮石斛
DoANS
和
UDP-
葡萄糖类黄酮
-3-
O
-
葡萄糖基转移酶
DoUFGT
是合成花色苷的关键基因,可影响飞燕草素和矢车菊素的累积,使茎表现出红色。研究表明,不同品种色泽差异与花色素苷生物合成酶基因转录水平差异有关,如胡萝卜紫色品种中
PAL3
/
PAL4
,
CA4H1
和
4CL1
的转录水平高于非紫色品种;且
CHS1
,
CHI1
,
F3H1
,
F3'H1
,
DFR1
和
LDOX1/LDOX2
仅在紫色品种主根中高表达,而在非紫色品种主根中不表达或很少表达。由于矢车菊苷
3-
葡萄糖苷基因结构变异,金银花中
DFR
,
LDOX
,
BZ1
和
GT1
的转录水平低于红金银花。
花色素苷生物合成酶基因转录水平的变异受到转录复合体
MBW(MYB-bHLH-WD40)
的协同调控。其中
R2R3-MYB
转录因子在花色素苷生物合成的转录调控中起关键作用,如银杏叶
GbMYBFL
的转录水平与其类黄酮含量呈正相关,且
GbMYBFL
过表达可诱导类黄酮和花青素的积累。灰毡毛忍冬叶片
MYB12
,
MYB75
,
bHLH113
,
TTG1
也被认为是调节黄酮苷生物合成的关键基因。淫羊藿
EsMYBA1
和
EsAN2
参与花色素苷的调控,但二者的表达组织特异性不同,其中
EsMYBA1
主要在叶片中表达,而
EsAN2
主要在花和花蕾中表达;且在烟草中过表达
EsAN2
可提高查尔酮合酶(
CHS
)、查尔酮异构酶(
CHI
)和花青素合酶(
ANS
)以及
bHLH
转录因子
NtAn1a
,
NtAn1b
的转录水平。从黑果枸杞和宁夏枸杞中分别克隆
MYB
转录因子
LrAN2
,
LbAN2
,并发现
2
个基因在烟草中过表达可诱导花色苷的生物合成,且
LbAN2
转基因品系叶片中的花色苷含量低于
LrAN2
,暗示
AN2
功能多样性可能是两种枸杞花色苷含量差异的原因。
AP2/ERF
转录因子可以调控丹参酮和丹参酚酸的累积,研究表明过表达
SmERF115
可使丹参毛状根中酚酸的产量增加,同时降低丹参酮含量;而沉默
SmERF115
可提高丹参酮含量。植物光形态建成的负调控因子
COP
,
PbCOP1.1
可以显著抑制花青素生物合成基因
CHI
,
DFR
,
UFGT2
,
bHLH3
,
HY5
,
GST
的表达,从而抑制花青素的积累。
2.2
吲哚类物质
一般认为吲哚合成途径主要通过莽草酸途径,即莽草酸先形成分支酸,分支酸在邻氨基苯甲酸合酶作用下形成邻氨基苯甲酸后,在吲哚
-3-
甘油磷酸合成酶作用下形成吲哚
-3-
甘油磷酸,然后吲哚
-3-
甘油磷酸在色氨酸合成酶作用下形成吲哚;吲哚经细胞色素
P450
单加氧化酶氧化为
2-
羟基吲哚和
3-
羟基吲哚,吲哚酚加氧聚合成靛蓝、靛玉红或衍生为其他吲哚类物质。
靛蓝、靛玉红属同分异构体,是中药青黛和板蓝根中含量最高的活性成分,对治疗慢性粒细胞白血病具有显著疗效。植物靛类生物合成的关键步骤是吲哚
-3-
甘油磷酸裂解酶(
IGL
)生产吲哚,再在吲哚合酶基因(
PtINS
)的参与下形成靛蓝。
PtUGT1
是一种
UDP-
葡糖基转移酶基因,利用葡萄糖基作为生物化学保护基团,抑制吲哚酚形成靛苷。从细菌中分离的蛋白酶如
CYP102A_scat
能够催化吲哚分子的细胞内羟基化,促进靛蓝的合成,当另外添加靛蓝和靛玉红生物合成途径的前体物质色氨酸时,靛蓝和靛玉红的产量也会提高。经过设计的人造酶
F43YMb
和
F43Y/H64DMb
可产生具有高达
80%
的化学选择性的靛蓝产物。
Met446
基因座中的突变,使戊糖磷酸途径成为葡萄糖的主要代谢途径,为吲哚的酶促氧化提供了更多的辅因子
NADPH
,靛玉红的效率进一步提高。
2.3
类胡萝卜素等萜类化合物
类胡萝卜素是一种天然色素,广泛存在于动物、高等植物、真菌、藻类。目前已知的类胡萝卜素主要分为两大类,一是含氧类胡萝卜素,如玉米黄质、堇菜黄质、新黄质和叶黄质等,被称为叶黄素类;二是不含氧的类胡萝卜素,如
α
-
胡萝卜素、
β
-
胡萝卜素和番茄红素,被称为胡萝卜素。其生物合成途径主要包括两步,首先是异戊烯焦磷酸(
IPP
)在
IPP
异构酶(
IPPI
)催化下生成二甲基丙烯基二磷酸(
DMAPP
),
DMAPP
在牻牛儿基牻牛儿焦磷酸(
GGPP
)合成酶(
GGPS
)作用下与
3
分子
IPP
缩合,分别生成牻牛儿焦磷酸(
GPP
)、法呢基焦磷酸(
FPP
)、
GGPP
。其中
FPP
是萜类衍生物如长醇、甾醇、倍半萜等和类胡萝卜素的合成前体物质,
GGPP
则可被进一步催化生成八氢番茄红素、叶绿素、维生素
A
、赤霉素、磷酸甘油醛、质体醌、泛醌等。第二步,由八氢番茄红素合成酶(
PSY
)催化的
2
分子
GGPP
,形成八氢番茄红素,八氢番茄红素经过八氢番茄红素脱氢酶(
PDS
)和六氢番茄红素脱氢酶(
ZDS
)的
4
步连续脱氢形成番茄红素,在
2
种番茄红素环化酶(
LCYb
、
LCYe
)作用下形成胡萝卜素,胡萝卜素经过羟化酶和加氧酶的作用形成种类多样的类胡萝卜素及其他下游代谢产物。
金银花中类胡萝卜素裂解双加氧酶
LjCCD4
和
LjCCD1b
在花着色中发挥重要作用。苦瓜
McGGPPS1
,
McGGPPS2
,
McPSY
,
McPDS
,
McZDS
,
McLCYB
,
McLCYE1
,
McLCYE2
,
McXHB
和
McZEP
基因参与了类胡萝卜素积累,其基因表达量在不同器官及果实不同成熟阶段也存在差异。枸杞类胡萝卜素生物合成基因表达水平也呈现组织特异性,即
LcIPI
在花中表达最高,
LcPSY
,
LcPDS
,
LcZDS
和
LcCRTISO
在成熟果实中表达最高,
LcLYCE
在叶片中表达最高,且关键酶基因
LcPDS
,
LcZDS
和
LcCRTISO
还具有提高类胡萝卜素含量和植物耐盐性的潜力。
CCS
,
PSY
,
LCY-B
和
CRT-Z
等也是辣椒红素合成的关键基因,其表达模式的多样性导致了辣椒果实颜色的多样性。当沉默单个基因时,红色品种的辣椒果实呈橙色或黄色;当几个基因同时沉默时,果实不能显示正常的红色;当单个或几个基因同时被沉默时,与辣椒红素合成相关的
β
-
胡萝卜素、
β
-
隐黄素或玉米黄质含量呈不同程度的降低,而辣椒红素含量降低最为显著。加利福尼亚罂粟中
PDS
,
ZDS
,
βOH
和
ZEP
基因的沉默可使其总色素含量降低
75
~
90%
,并改变了花瓣的颜色。在自然界中,环境因素在很大程度上影响植物类胡萝卜素的组成,如
HHP
处理(
50000
~
400000kPa
,
3
~
60min
,
4℃
下储存)番木瓜果实后,其八氢番茄红素去饱和酶转录水平增加,类胡萝卜素前体和类胡萝卜素含量也增加,其中番茄红素含量增加了
11
倍。
作为决定药材质量的物质基础之一,药材中的花色素苷、吲哚类物质、类胡萝卜素等色素成分的合成与调控受到光照、紫外线辐射、采收时期等外界环境因子共同作用的影响。
3.1
光照
光照是影响花色素苷合成的重要环境因子,通过影响光合作用可促进糖、苯丙氨酸等花色素苷前体物质的合成,同时光的信号转导也可调控花色素苷生物合成基因及其相关转录因子的表达,进而促进花色素苷的积累。在光照条件下,光受体可通过与
COP1
互作,抑制
COP1
的负向调控功能,使其下游转录因子
HY5
可通过
MYB
激活花色素苷的生物合成,从而促进花色素苷的积累;在黑暗条件下,
COP1
可通过泛素化降解
HY5
,抑制花色素苷的积累。
HY5
还可通过抑制花色素苷负向调控因子
MYBL2
的表达,提高
MBW
复合体的稳定性,进而促进花色素苷下游生物合成基因的表达,引起花色素苷积累。紫外辐射如
UV-A
,
UV-B
,
UV-C
等也能通过诱导丙苯氨酸途径
VcPAL
,
VcUFGT
等或抑制
VcDFR
等基因的转录,促使花青苷的积累。
UV-B
处理可促进黄花蒿
3-






