
高等植物SA的生物合成途径一直是引人注目的科学问题,介于第一步CA-ISC的异构化反应在细菌和植物中高度保守,研究者们普遍认为植物中存在类似细菌IPLs的酶催化第二步反应,但一直苦寻无果。
2019年4月5日,
BioRxiv
杂志在线刊登了了
加拿大英属哥伦比亚大学
张跃林
实验室
和
德国哥廷根大学
Ivo Feussner
实验室
题为 “
From isochorismate to salicylate: a new reaction mechanism for salicylic acid biosynthesis
”的研究论文。该研究表明拟南芥PBS3蛋白作为氨基转移酶催化异分支酸(ISC)形成异分支酸-谷氨酸加合物(ISC-9-Glu),随后ISC-9-Glu自发分解形成水杨酸(SA),打通了SA合成途径的最后一环。
值得一提的是,在该文发表了2天后,
BioRxiv
杂志在线刊登了来自
美国博德研究所和麻省理工学院
Jing-Ke Weng
实验室
题为“PBS3 and EPS1 complete salicylic acid biosynthesis from isochorismate in
Arabidopsis
”的文章。
该研究同样表明拟南芥PBS3蛋白是异分支酸(ISC)转化为水杨酸(SA)的关键催化因子,同时还发现EPS1能够以酶促方式促进PBS3蛋白催化产物ISC-9-Glu分解形成SA。注:
详细见
我们公众号iPlants在今天的第二条推送。
因此,
这两项研究都证实PBS3蛋白正是SA生物合成途径所缺失的最后一环,
两篇研究的切人点和论证思路都高度相似,结果相互印证,值得学习借鉴!
水杨酸(salicylic acid, SA)在调控植物免疫和生长方面具有广泛作用。前期的研究表明,病原菌诱导的SA生物合成主要依赖于叶绿体途径。现有的模型认为,在叶绿体中,
分支酸
(chorismic acid, CA ) 首先在
异分支酸合酶
( isochorismate synthase 1 , ICS1) 的催化下形成
异分支酸(ISC)
,接着ISC以未知的方式转化为SA,最终通过转运体蛋白 EDS5 转运至胞质。在细菌中也存在类似的CA依赖的SA合成途径,既可以通过
双功能的SA合酶 (SA synthase,SAS)
一步完成,也可以先异构化形成ISC,再通过异分支酸丙酮酸裂合酶(isochorismate pyruvate lyase, IPL)形成SA (见下图)。遗憾的是,
拟南芥ICS1并不具备SAS的双重催化功能,且在植物基因组中也没有发现IPLs的存在,所以植物ISC-SA途径的生化机制至今仍是一个谜团!
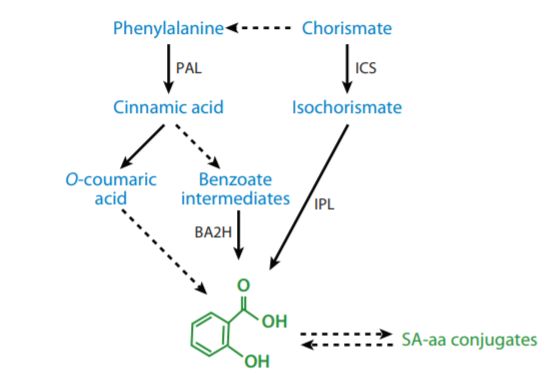
图. SA生物合成途径的简化示意图(Annu. Rev. Phytopathol. 2009. 47:177–206)
在该研究中,作者首先注意到
拟南芥
中和催化茉莉酸-亮氨酸加合物(JA-Ile)形成的氨基转移酶AtGH3.11(JAR1)同属
GH3蛋白家族的
PBS3
突变体中SA的含量减少,但PBS3蛋白并不能以SA作为底物,因而
pbs3
突变体中SA的含量减少应该另有原因。
为了进一步研究PBS3蛋白的功能,作者首先通过杂交将
pbs3
突变引入自发免疫突变体
snc2-1D/ npr1-1
,发现和之前报道的
snc2-1D/npr1-1/eds5-3
三突变体类似,不能回复双突的矮化表型。代谢物指纹分析显示,PBS3和EDS5突变都能显著降低
snc2-1D/npr1-1
突变体中SA及其衍生物的含量 (Fig. 1)。有趣的是,
snc2-1D/npr1-1/pbs3
突变体相比
snc2-1D/npr1-1/eds5-3
突变体有SA前体 ISC的累积,因而推测 ISC可能是PBS3蛋白的底物。
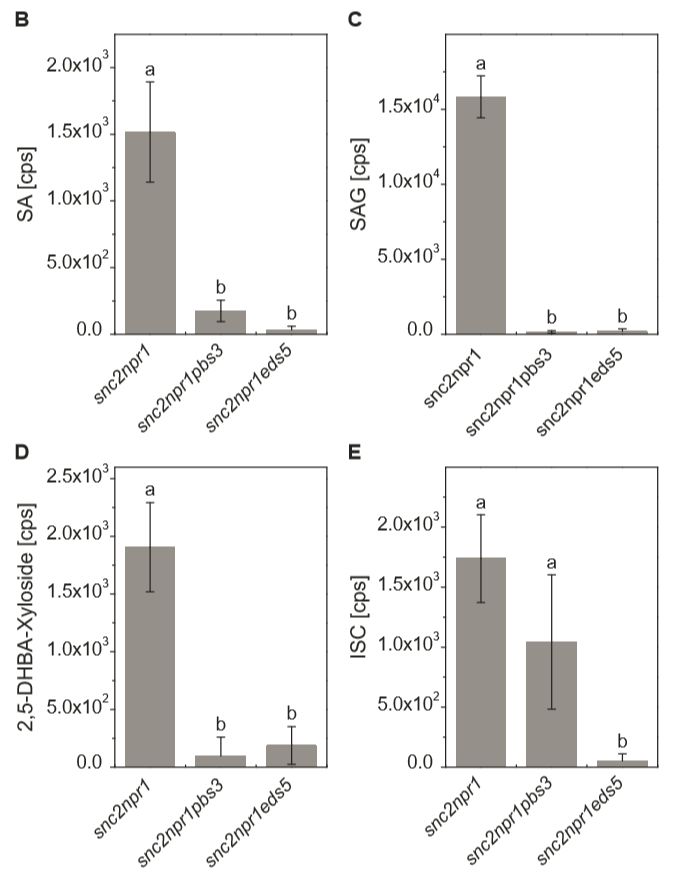
Figure 1. 拟南芥突变体SA相关代谢物指纹分析
随后,作者通过异源表达系统纯化了PBS3蛋白,在体外测试了其催化活性。质谱分析显示PBS3蛋白既可以催化CA的 C7和 C9位的谷氨酸化(CA-7-Glu, CA-9-Glu),也可以以ISC为底物生成ISC-9-Glu,且表现出对ISC的底物偏好性。意外的是,在催化产物中也检测到了SA的存在。由于此前有研究表明在溶液中ISC自身可以缓慢转变为SA,所以作者检测了在研究体系中SA的生成是否依赖于PBS3的催化反应?研究发现无论是去除Glu、ATP或PBS3蛋白,SA的生成速率相比完全反应时都要显著降低,
暗示ISC-9-Glu的形成可以强烈地促进ISC骨架的分解,进而导致SA的累积
(Fig. 2)。反应动力学分析显示,PBS3蛋白对ISC有很高的亲和力和催化效率。基于PBS3-SA-AMP晶体结构的数据模拟也发现ISC可以适配PBS3的催化活性中心,且ISC的C9位氨基酸侧链基团和AMP分子的磷酸基团在空间上紧邻。
基于以上证据,作者认为PBS3蛋白可能直接参与SA的生物合成。
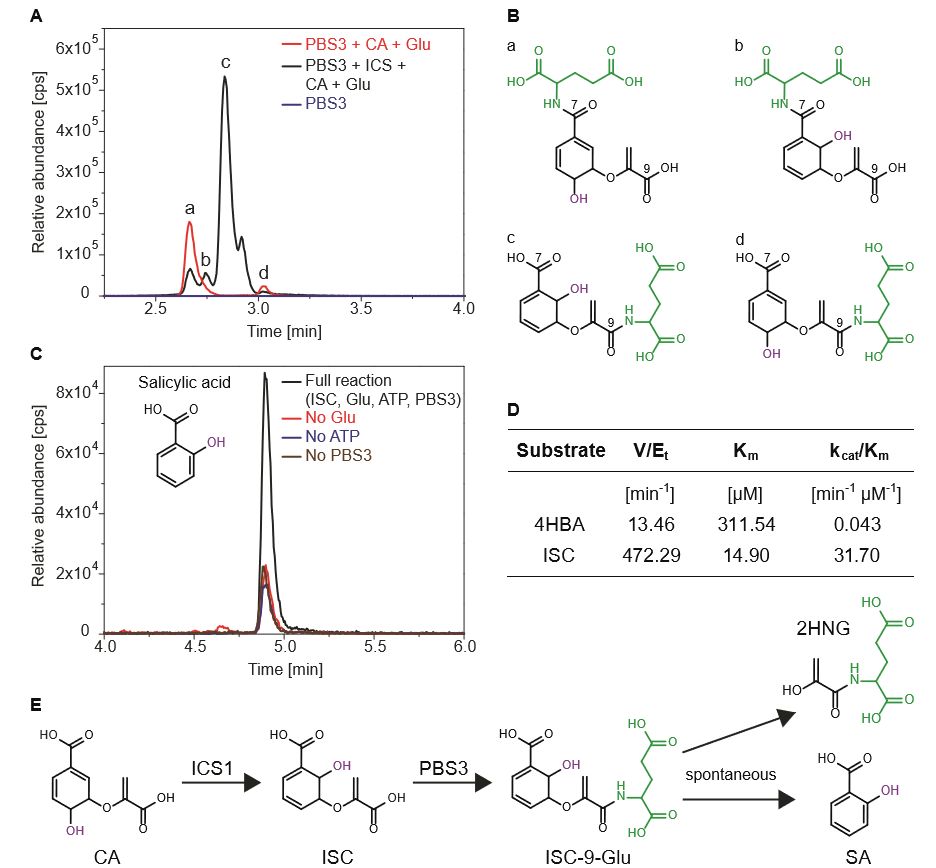
Figure 2. 体外实验中PBS3催化ISC-9-Glu生成及
ISC-9-Glu自发分解生成SA的反应特征
为了进一步验证ISC-9-Glu的自发分解途径在植物中是否存在,作者检测了
snc2-1D/npr1-1
突变体的代谢物,发现确实能检测到 ISC-9-Glu分解产物
SA和2HNG
,但
snc2-1D/npr1-1/pbs3
突变体和
snc2-1D/npr1-1/eds5-3
突变体中,SA和2HNG几乎检测不到(Fig. 3A, B)。此外,通过串联质谱进一步确认体内和体外来源的 ISC-9-Glu具有一致的二级图谱,因而认为
ISC-9-Glu是植物体内是SA的直接前体,通过快速的自发分解生成
SA
。
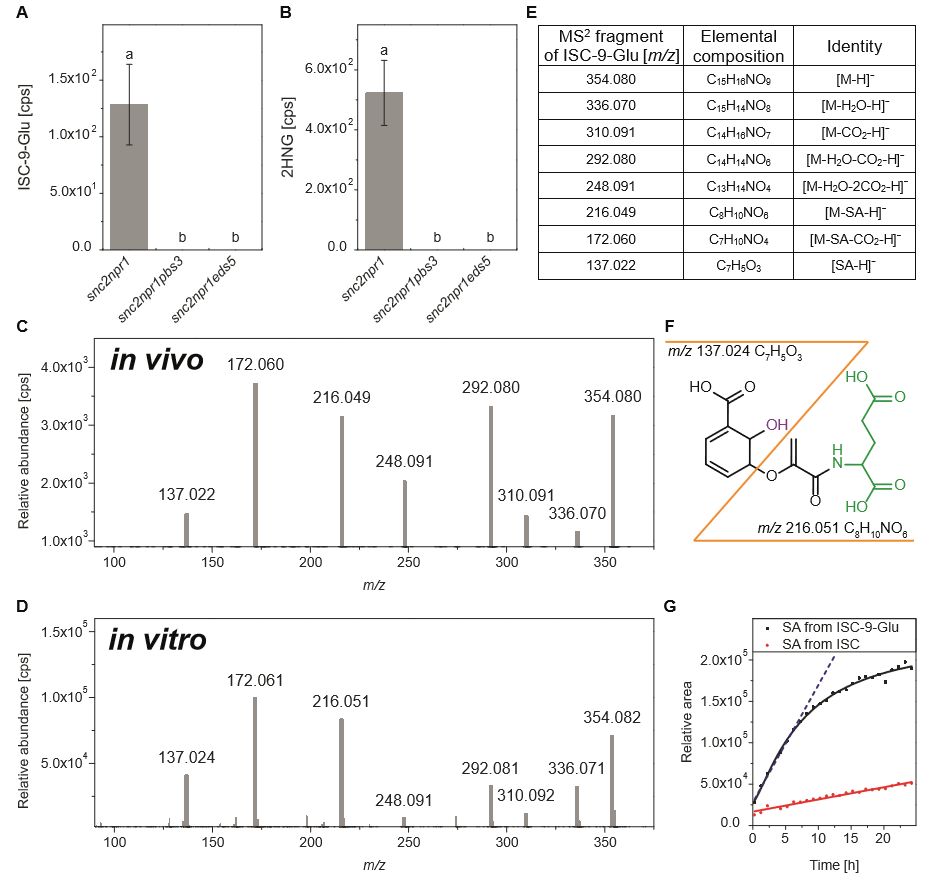
Figure 3. 质谱分析证实
ISC-9-Glu自发分解产物SA和2HNG的存在
接下来,作者分析了ISC-9-Glu分解过程的分子动力学特征,发现ISC-9-Glu在反应开始6h内以线性速率发生解离,在20h达到平衡(Fig. 3G)。分子模拟显示 ISC-9-Glu可以形成三个氢键,其中谷氨酸骨架的氨基氢原子和醚键氧原子形成的氢键拉近了两个官能团的距离,能够促进质子的转移,引发ISC碳环的芳构化,通过单分子消除反应生成SA和2HNG (Fig. 4)。ISC-9-Glu通过分子内氢键作用自发形成类似于细菌IPL催化ISC形成的活化过渡态,在反应特征上具有保守性。
上述SA通过非酶促反应生成的化学特征也可以部分解释病原菌诱导的SA累积为何迟滞于ICS1, PBS3和EDS5基因的表达。
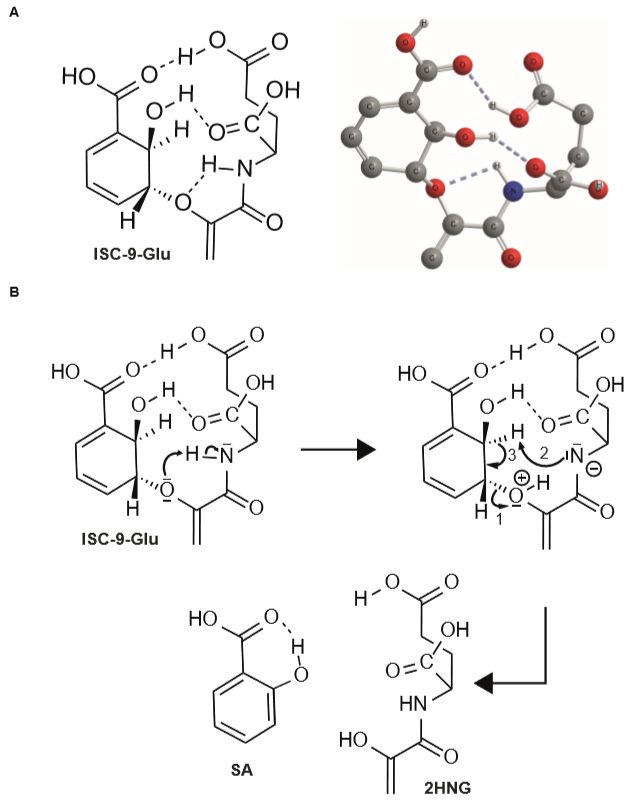
Figure 4. ISC-9-Glu自发分解过程的分子模型
此前的模型认为SA 的全合成发生在叶绿体中,并通过EDS5将SA转运至胞质。然而,无论是基于序列特征的软件预测还是GH3家族其它蛋白的研究,
都支持PBS3是一个细胞质蛋白
。为了验证PBS3蛋白的定位,作者首先通过在烟草和拟南芥叶片中瞬时表达PBS3-YFP融合蛋白(遗传回补实验证实该融合蛋白具有功能),但没有观察到荧光信号。为此,作者转而将ICS1的叶绿体转运肽融合在PBS3-YFP的N端,构建了chloroPBS3-YFP,并和ICS1-CFP共转拟南芥叶片,发现PBS3和ICS1在叶绿体有共定位。 将chloroPBS3与ICS1共转eds5-3突变体,发现可以恢复突变体的SA累积缺陷(Fig. 5B),而PBS3本身或者与ICS1共转都不能恢复,
暗示EDS5的作用是将ICS1催化形成的ISC转运出叶绿体,在胞质中由PBS3转化为ISC-9-Glu
,这也解释了为何eds5-3突变体中SA含量减少。

Figure 5. A-B, PBS3和ICS1共定位表达驱动SA合成;C, SA生物合成的模型
综上所述,该研究提供了充分的生化和遗传证据,证实SA生物合成的叶绿体途径由ICS1、EDS5 和PBS3接力完成,首先在叶绿体中CA通过ICS1异构化形成ISC,经EDS5转运出叶绿体,在胞质中由PBS3催化形成ISC-9-Glu,随后ISC-9-Glu自发分解生成SA (见下图)。
由于PBS3的同源蛋白在植物中广泛存在,本研究揭示的机制很可能是高等植物SA合成的基本途径。
编者注:
高等植物SA的生物合成途径一直是引人注目的科学问题,介于第一步CA-ISC的异构化反应在细菌和植物中高度保守,研究者们普遍认为植物中存在类似细菌IPLs的酶催化第二步反应,但一直苦寻无果。本研究发现实际上
ISC是通过PBS3形成
ISC-9-Glu,随后自发分解生成SA,该自发过程模拟了被
IPLs活化的
ISC状态。高等植物进化出的这一步SA非酶促合成步骤有效地避免了
IPLs反应专一性差的问题,可以实现SA的快速、定向合成。此外,
PBS3蛋白很早就被报导为植物免疫正调节因子,影响病原菌诱导的SA累积,但十几年来一直不清楚其作用机理。受到同源蛋白JAR1功能研究的误导,
PBS3蛋白的底物曾被认为是SA,但很快被生化证据所否认。尽管近期有研究报导
PBS3蛋白可以通过蛋白-蛋白相互作用参与SA信号的调节,但其作为酶的功能却一直未被揭示。本研究的发现给人以拨云见日之感,实是领域的重大突破,看文章的行文格式完全是
Science
的风格,期待
!
论文链接或点击【阅读原文】:
https://www.biorxiv.org/content/biorxiv/early/2019/04/05/600692.full.pdf
注:由于该文报道的论文还在预印杂志上,故未经允许谢绝其他公众号转载,以避免产生与本文不实的信息!
微信加群
iPlants组建了7个植物科学研究交流群,汇聚了一批全球各地的从事植物科学研究的研究生和PI。欢迎从事植物科学相关研究的同学和老师加入我们,一起讨论学术和梦想。
温馨提示:进群请备注一下(格式如学校+专业+姓名,如果是PI/教授,请
注明是
PI/教授/副教授/副研究员/博后
,否则就直接
默认为在读研究生
,谢谢
)。
加小编微信号(ID:
iplants
)
或长按下面二维码
时,请注明学校和专业,非诚勿扰。

iPlants,专注植物最前沿。
投稿、商务合作、转载开白名单等事宜请联系微信ID:
iplants
或邮箱:
[email protected]
觉得本文好看,请点这里↓





