写在前面的话
不论大家是做什么方向,人或者某个物种,其基因组的获取对于加速科研产出意义重大,但是获取一个物种的基因组的过程不仅仅包括测序,也包括组装,组装部分十分体现生信分析者的水平,今天跟大家聊一下组装的前世今生。
简介
从1977年Sanger发明“双脱氧链终止法”DNA测序技术起,基因组神秘的面纱一点一点的被揭露,从小至几千碱基的噬菌体基因组到数百万碱基的细菌基因组,再到三十亿碱基的人类基因组,每一步都值得记录在人类探索自然、认识自身的篇章中。而这些成果背后的重要一环——基因组组装,无疑是一个在研究中足够”美”的问题:既足够简明,可用短短的一段话来描述;又足够深刻,值得数十年的持续研究。小编今天和大家分享分享基因组组装的历史以及前沿的发展。
Part 1.白云生处

上面是计算机学家Staden关于序列拼接的描述,从中可以引申出我们如今经常使用的几个术语:reads/overlap/contig。对于序列拼接的概念Staden进行了简明的定义:通过读取片段(reads)间的连接关系(overlap)构建出更长的连续性片段(contig)。
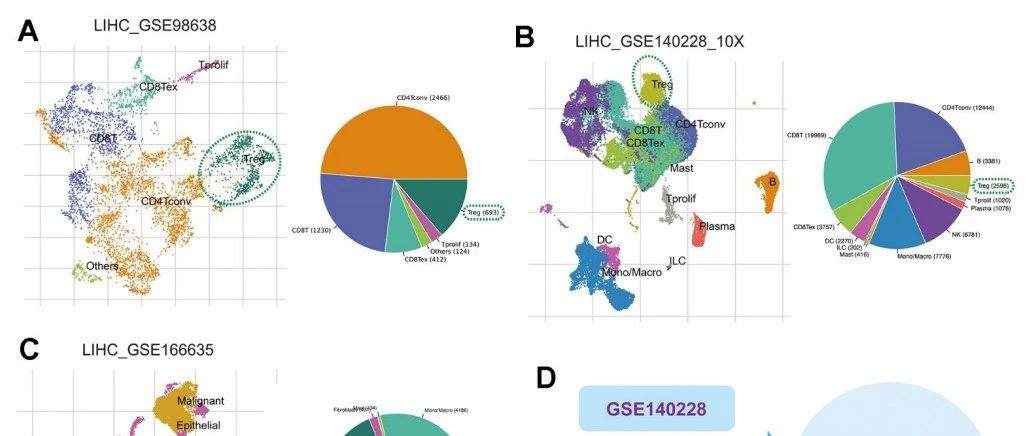
更进一步的组装研究中,序列拼接问题被转化为图论中的路径寻找问题:以点(node)代表测序序列,以边(edge)代表连接关系,以路径(path)代表的图上点的定向行走(walk)。这里面,最有代表性的两种构图方式即string图和de Brujin图。下面奉上两张小编收藏多年的图,非常好的阐述了string图和de Brujin图在基因组组装中的应用原理。
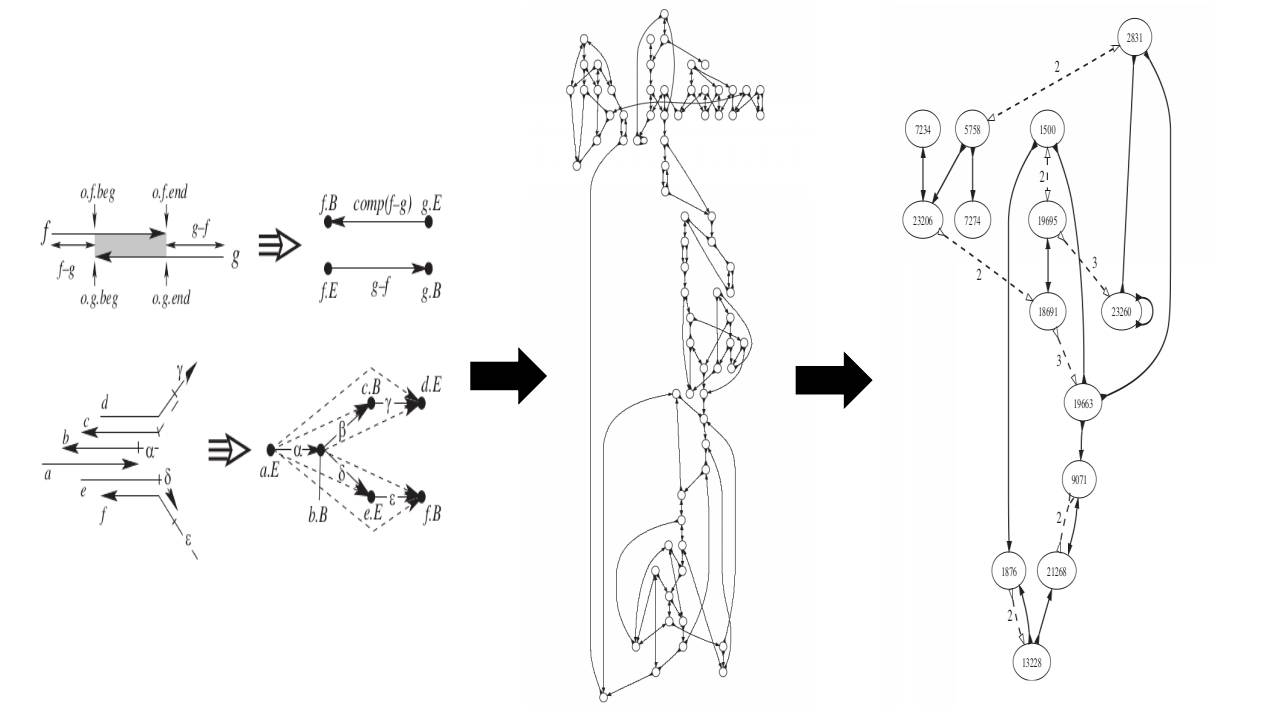
图1. Strings Graph in genome assembly[2]
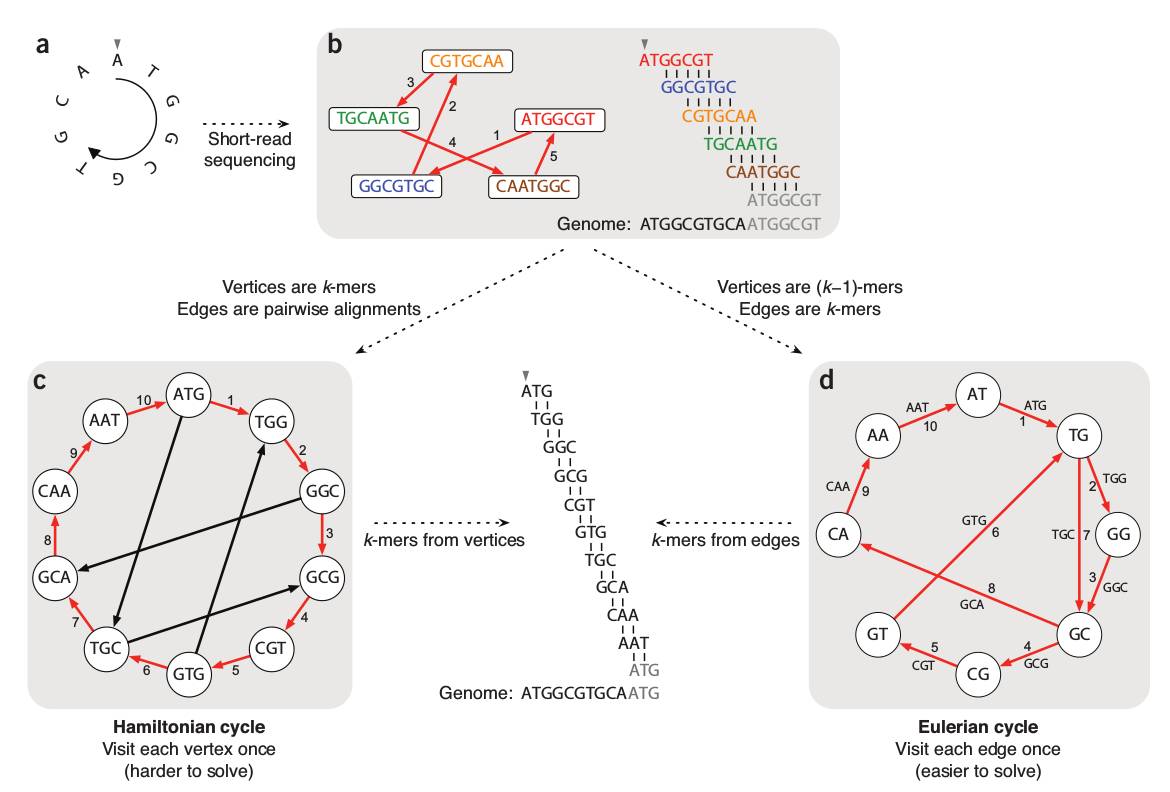
图2. De Brujin Graph in genome assembly[3]
Part 2.一往而深
提起基因组,最广为人知的应该是人类基因组计划了,2001年公布的人类基因组是这一计划的里程碑事件。其中,大放光彩的Celera Assembler也成为基因组组装的”初代机“[4],whole genome shotgun的测序策略结合Overlap Layout Consensus的组装策略,攻克了基因组学研究上的第一座高峰。
但是一代测序由于高昂的测序成本以及较低的测序通量,限制了其在更多、更大规模的基因组学研究中的应用。随着二代高通量测序的应运而生,全基因组测序才成为科研人员广泛使用的工具。以2005年出现的454测序仪和2008年出现的illumina测序仪为代表,短读长、高通量的测序数据成为主流。而对于基因组组装而言,与之而来的却是短至几十碱基的测序片段带来的拼接困境。
为此,研究人员发明了不同的文库构建方法,以及改变了序列拼接的算法。高深度+多文库的双端测序策略结合de Brujin图的组装策略,成为新一代的组装标杆。在这一风起云涌的时代,华大基因以其SOAPdenovo[5],以快打慢,打下了一片大大的江山(大雾)。
所谓一代版本一代神,虽然通过二代测序绘制了多物种的基因组草图,但整体的连续性和完整性上仍存在较大不足。随着三代单分子测序技术的出现,又再次焕发了OLC组装策略的新春。基于Celera Assembler,研究人员适应三代测序数据形成了Hierarchical Genome Assembly Process(HGAP)的先纠错再组装的策略[6]。而二代测序并没有因此退出组装舞台,采用巧妙的文库构建方法如全基因组染色体构象捕获测序技术(Hi-C)、在DNA片段上加入高通量的barcode标签测序技术(10X)等,能够进一步对基因组进行完善升级,甚至使组装结果达到染色体水平。
说起来,小编最初接触组装时深入研究的就是Celera Assembler,当时还是三代测序出现之初的7.0版本,见证了诸多版本的更新,不得不说,开发人员确实是一往而深(大雾),当为吾辈楷模。
Part 3.沧海云帆
组装的终极目标是得到一个没有间隙(gap)的、单倍体精度的组装结果,但是目前为止,还没有一个高等动植物的基因组实现这样的目标。即使是研究最完善的人类基因组,目前仍存在800余个gaps。但是这个目标的实现距离我们已经是触目可及了:测序技术不断发展,一代、二代、三代、光学等数据优势互补,使组装如虎添翼;建库方法不断改进,Hi-C、10X等方法画龙点睛,助组装锦上添花。
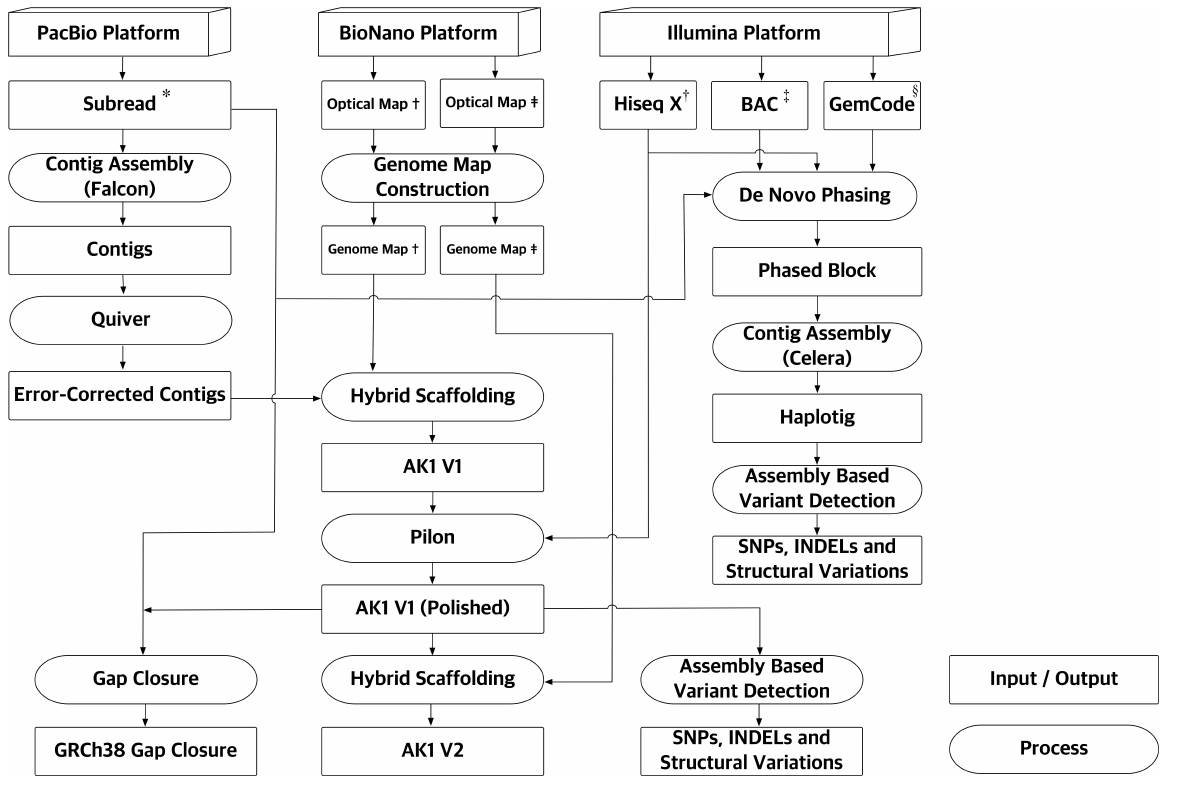
图3. 测序组装策略[7]
如果问小编,组装的未来是什么呢?小编最想看到的是没有组装!一条DNA从头测到尾,0 gap,不组装,测出即用。有可能实现吗?让我们一起拭目以待吧。